Personality traits modulate subcortical and cortical vestibular and anxiety responses to sound-evoked otolithic receptor stimulation Iole Indovina a,b, ⁎, Roberta Riccelli c , Jeffrey P. Staab d , Francesco Lacquaniti a,b,e , Luca Passamonti f a Laboratory of Neuromotor Physiology, IRCCS Santa Lucia Foundation, 00179, Rome, Italy b Centre of Space BioMedicine, University of Rome Tor Vergata, 00173, Rome, Italy c Department of Medical and Surgical Sciences, University “Magna Graecia”, Catanzaro, Italy d Department of Psychiatry and Psychology, Mayo Clinic, Rochester, MN, USA e Department of Systems Medicine, Neuroscience Section, University of Rome Tor Vergata, 00133 Rome, Italy f Institute of Bioimaging and Molecular Physiology, National Research Council 88100, Catanzaro, Italy abstract article info Article history: Received 20 March 2014 Received in revised form 31 July 2014 Accepted 5 September 2014 Keywords: Anxiety Neuroticism Introversion Sound-evoked vestibular stimulation fMRI Chronic subjective dizziness Objective: Strong links between anxiety, space-motion perception, and vestibular symptoms have been recog- nized for decades. These connections may extend to anxiety-related personality traits. Psychophysical studies showed that high trait anxiety affected postural control and visual scanning strategies under stress. Neuroticism and introversion were identified as risk factors for chronic subjective dizziness (CSD), a common psychosomatic syndrome. This study examined possible relationships between personality traits and activity in brain vestibular networks for the first time using functional magnetic resonance imaging (fMRI). Methods: Twenty-six right-handed healthy individuals underwent fMRI during sound-evoked vestibular stimu- lation. Regional brain activity and functional connectivity measures were correlated with personality traits of the Five Factor Model (neuroticism, extraversion-introversion, openness, agreeableness, consciousness). Results: Neuroticism correlated positively with activity in the pons, vestibulo-cerebellum, and para-striate cortex, and negatively with activity in the supra-marginal gyrus. Neuroticism also correlated positively with connectivity between pons and amygdala, vestibulo-cerebellum and amygdala, inferior frontal gyrus and supra-marginal gyrus, and inferior frontal gyrus and para-striate cortex. Introversion correlated positively with amygdala activity and negatively with connectivity between amygdala and inferior frontal gyrus. Conclusions: Neuroticism and introversion correlated with activity and connectivity in cortical and subcortical vestibular, visual, and anxiety systems during vestibular stimulation. These personality-related changes in brain activity may represent neural correlates of threat sensitivity in posture and gaze control mechanisms in normal individuals. They also may reflect risk factors for anxiety-related morbidity in patients with vestibular disorders, including previously observed associations of neuroticism and introversion with CSD. © 2014 Elsevier Inc. All rights reserved. Introduction Anxiety influences postural control and locomotion in health and disease, though neural mechanisms responsible for these effects are incompletely understood. In the 1870s, publication of the original defi- nition of agoraphobia (i.e., spatial disorientation and fear in busy town squares) sparked debates about interactions among anxiety, spatial orientation, and gait [1]. A century later, these interactions were inves- tigated in clinical and laboratory studies [1,2]. Healthy people, when positioned at height, were found to have a stiffer stance and experience greater state anxiety, lower balance confidence, and higher autonomic arousal [3–9]. Individuals with high trait anxiety stiffened their stance under modest stress that did not alter postural control of those with low trait anxiety [10]. Conversely, vestibular disorders were identified as potent triggers of secondary anxiety disorders [11–18] and elevated state anxiety was associated with lower functional status [19] and poorer treatment outcomes [19,20]. Animal studies provided evidence about important neural connec- tions between subcortical anxiety and vestibular systems [21–26]. In- formation about head motion is conveyed from vestibular labyrinths to vestibular nuclei, and then to pathways that control oculomotor and spinal reflexes, modulated by loops through the cerebellum. This information reaches systems that mediate affective responses via path- ways from vestibular nuclei through the pontine nuclei to the amygdala [2,21–23]. In the cortex, vestibular, visual, proprioceptive, and somato- sensory information about space and motion reach associative regions via the thalamus. In monkeys, the core vestibular cortex is called the parieto-insular vestibular cortex. The human homologue of this region includes sylvian and peri-sylvian areas at the interface between the posterior- and retro-insula, posterior superior temporal gyrus (STg) Journal of Psychosomatic Research 77 (2014) 391–400 ⁎ Corresponding author at: Santa Lucia Foundation, Via Ardeatina 306, 00179 Rome Italy. E-mail address: [email protected] (I. Indovina). http://dx.doi.org/10.1016/j.jpsychores.2014.09.005 0022-3999/© 2014 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Journal of Psychosomatic Research

Personality Traits Vestibulo Cohlear
Dec 11, 2015
personality
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Psychosomatic Research 77 (2014) 391–400
Contents lists available at ScienceDirect
Journal of Psychosomatic Research
Personality traits modulate subcortical and cortical vestibular andanxiety responses to sound-evoked otolithic receptor stimulation
Iole Indovina a,b,⁎, Roberta Riccelli c, Jeffrey P. Staab d, Francesco Lacquaniti a,b,e, Luca Passamonti f
a Laboratory of Neuromotor Physiology, IRCCS Santa Lucia Foundation, 00179, Rome, Italyb Centre of Space BioMedicine, University of Rome Tor Vergata, 00173, Rome, Italyc Department of Medical and Surgical Sciences, University “Magna Graecia”, Catanzaro, Italyd Department of Psychiatry and Psychology, Mayo Clinic, Rochester, MN, USAe Department of Systems Medicine, Neuroscience Section, University of Rome Tor Vergata, 00133 Rome, Italyf Institute of Bioimaging and Molecular Physiology, National Research Council 88100, Catanzaro, Italy
⁎ Corresponding author at: Santa Lucia Foundation, VItaly.
E-mail address: [email protected] (I. Indovina)
http://dx.doi.org/10.1016/j.jpsychores.2014.09.0050022-3999/© 2014 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 20 March 2014Received in revised form 31 July 2014Accepted 5 September 2014Keywords:AnxietyNeuroticismIntroversionSound-evoked vestibular stimulationfMRIChronic subjective dizziness
Objective: Strong links between anxiety, space-motion perception, and vestibular symptoms have been recog-nized for decades. These connections may extend to anxiety-related personality traits. Psychophysical studiesshowed that high trait anxiety affected postural control and visual scanning strategies under stress. Neuroticismand introversion were identified as risk factors for chronic subjective dizziness (CSD), a common psychosomaticsyndrome. This study examined possible relationships between personality traits and activity in brain vestibularnetworks for the first time using functional magnetic resonance imaging (fMRI).Methods: Twenty-six right-handed healthy individuals underwent fMRI during sound-evoked vestibular stimu-lation. Regional brain activity and functional connectivity measures were correlated with personality traits ofthe Five Factor Model (neuroticism, extraversion-introversion, openness, agreeableness, consciousness).Results:Neuroticism correlated positivelywith activity in the pons, vestibulo-cerebellum, and para-striate cortex,andnegativelywith activity in the supra-marginal gyrus. Neuroticism also correlated positivelywith connectivity
between pons and amygdala, vestibulo-cerebellum and amygdala, inferior frontal gyrus and supra-marginalgyrus, and inferior frontal gyrus and para-striate cortex. Introversion correlated positivelywith amygdala activityand negatively with connectivity between amygdala and inferior frontal gyrus.Conclusions: Neuroticism and introversion correlated with activity and connectivity in cortical and subcorticalvestibular, visual, and anxiety systems during vestibular stimulation. These personality-related changes inbrain activity may represent neural correlates of threat sensitivity in posture and gaze control mechanisms innormal individuals. They also may reflect risk factors for anxiety-related morbidity in patients with vestibulardisorders, including previously observed associations of neuroticism and introversion with CSD.© 2014 Elsevier Inc. All rights reserved.
Introduction
Anxiety influences postural control and locomotion in health anddisease, though neural mechanisms responsible for these effects areincompletely understood. In the 1870s, publication of the original defi-nition of agoraphobia (i.e., spatial disorientation and fear in busy townsquares) sparked debates about interactions among anxiety, spatialorientation, and gait [1]. A century later, these interactions were inves-tigated in clinical and laboratory studies [1,2]. Healthy people, whenpositioned at height, were found to have a stiffer stance and experiencegreater state anxiety, lower balance confidence, and higher autonomicarousal [3–9]. Individuals with high trait anxiety stiffened their stanceunder modest stress that did not alter postural control of those with
ia Ardeatina 306, 00179 Rome
.
low trait anxiety [10]. Conversely, vestibular disorders were identifiedas potent triggers of secondary anxiety disorders [11–18] and elevatedstate anxiety was associated with lower functional status [19] andpoorer treatment outcomes [19,20].
Animal studies provided evidence about important neural connec-tions between subcortical anxiety and vestibular systems [21–26]. In-formation about head motion is conveyed from vestibular labyrinthsto vestibular nuclei, and then to pathways that control oculomotorand spinal reflexes, modulated by loops through the cerebellum. Thisinformation reaches systems that mediate affective responses via path-ways from vestibular nuclei through the pontine nuclei to the amygdala[2,21–23]. In the cortex, vestibular, visual, proprioceptive, and somato-sensory information about space and motion reach associative regionsvia the thalamus. In monkeys, the core vestibular cortex is called theparieto-insular vestibular cortex. The human homologue of this regionincludes sylvian and peri-sylvian areas at the interface between theposterior- and retro-insula, posterior superior temporal gyrus (STg)

392 I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
and parietal operculum. Neuroimaging studies showed that humanvestibular cortical network includes the inferior parietal lobule [supra-marginal gyrus (SMg), temporo-parietal junction and ventral intra-parietal area], parts of the prefrontal cortex [e.g., the inferior frontalgyrus (IFg)], and the anterior cingulate cortex and hippocampus[27–33]. Vestibular and visual cortices interact in a reciprocally inhibito-ry manner. Vestibular stimuli activate vestibular cortex and suppressvisual cortical function, whereas visual motion stimuli do the opposite[34,35]. Thus, the human vestibular cortex is a multimodal processorthat integrates vestibular, visual, proprioceptive, somatosensory, andmotor information [36–38]. It also overlaps regions that process anxietyand affective responses, particularly in the posterior parietal, insular,hippocampal and prefrontal areas [25,39–41]. Thismay provide the cor-tical substrate for anxiety to influence vestibular information processingand for vestibular data to affect emotional states.
Researchers have developed increasingly sophisticated models ofvestibular–anxiety interactions at subcortical and cortical levels toexplain vestibular symptoms in patientswith primary anxiety disorders,and secondary anxiety disorders in patients with vestibular illnesses[2,21,23]. Thesemodelsmay extend to other conditionswith vestibular-anxiety interactions, such as height phobia and phobic postural vertigo[42,43] or chronic subjective dizziness (CSD) [2,44,45]. The latter twoare similar conditions that manifest with chronic non-vertiginous dizzi-ness or unsteadiness exacerbated by upright posture, head motion, andexposure to complex or moving visual stimuli, usually triggered byacute vestibular or balance disorders. They occupy a place in clinicalneurotology akin to irritable bowel syndrome in gastroenterology(i.e., independent psychosomatic conditions that occur with or withoutmedical or psychiatric comorbidity). A pre-existing anxiety diathesismay increase the risk of developing CSD [18,46], add chronic psychiatriccomorbidity [18,46], and reduce treatment response [47]. Personalitytraits of neuroticism and introversion were more closely associatedwith CSD than with comorbid neurotologic disorders producing similarlevels of vestibular and psychological symptoms [48]. In contrast,resilience, life satisfaction, and strong sense of coherence predictedlower rates of persistent dizziness after acute vestibular illnesses [49].Thus, personality traits associated with anxiety, specifically neuroticismand introversion may be important factors for anxiety-mediated vestib-ular conditions.
This untested hypothesis rests on the supposition that neurotic,introverted people compared to non-neurotic, extraverted individualshave: (1) central vestibular systems that are more reactive to vestibularstimuli, (2) anxiety systems that are more reactive to vestibular stimuli,or (3) central vestibular systems that reactmore strongly to inputs fromanxiety systems during vestibular stimulation. To investigate thesepossible mechanisms, we examined correlations between personalitytraits of the Five Factor Model (neuroticism, extraversion–introversion,agreeableness, openness, and conscientiousness) measured by theRevised NEO Personality Inventory (NEO-PI-R) [50] and brain activityand connectivity within vestibular and anxiety networks using func-tional magnetic resonance imaging (fMRI) in healthy volunteers stimu-lated with short tone bursts (STB) at 100 dB SPL. The STB procedure isknown to activate primary afferents in sacculi of the labyrinths andevoke robust responses in vestibular cortical areas [51–53]. By usinghealthy and drug-free volunteers imagedwhile in a comfortably relaxedstate, we focused on inherent relationships among personality traitsand brain activity during vestibular stimulation in subjects who werefree of potential confounds of high state anxiety, vestibular or anxietydisorders, and medications.
We predicted that higher neuroticism and lower extraversion scores(i.e., introversion) would correlate with increased reactivity to STBstimuli in subcortical and cortical vestibular and anxiety regions andgreater vestibular–anxiety system connectivity, thereby identifyingpotential neural substrates for clinical vestibular conditions rangingfrom comorbid anxiety in vestibular illnesses to CSD and phobias ofheights [2].
Participants and methods
Participants
Twenty-six healthy volunteers (14 females, mean age = 32.4 ±7.3 years) gave written informed consent to participate in this study,which was approved by the local Research Ethics Committee accordingto the Helsinki declaration (http://www.wma.net/en/30publications/10policies/b3/). Participants were systematically screened to excludemigraine, chronicmedical illnesses, pregnancy,medication use, smoking,or history of head injury. The MINI International NeuropsychiatricInventory was used to exclude those with past or present psychiatricdisorders [54]. All participants were right-handed, as assessed via theEdinburgh Handedness Inventory [55].
To measure personality traits of the Five Factor model, all partici-pants completed a computerized version of the Italian translation ofthe NEO-PI-R questionnaire [50]. Standardized T-scores for each per-sonality factor were calculated via a script written in SPSS (StatisticalPackage for Social Sciences, http://www.spss.it/) using combinedsex-norms reported in the NEO-PI-R manual.
fMRI task
Experimental stimuli were administered via piezo-electricMRI com-patible headphones (NordicNeuroLab,http://www.nordicneurolab.com/Products_and_Solutions/fMRI_Hardware/AudioSystem.aspx). Pre-vious studies demonstrated that unilateral STB stimuli at 100 dBsound pressure level (SPL) generate vestibular evokedmotor potentials(VEMPs) in sternocleidomastoid muscles ipsi- and contralateral to thestimulated ear, a response used in clinical practice to assess saccularfunction [51,53,56–58]. STB 100 dB SPL also evoke robust responses investibular cortical regions [51,53] (see also Supplementary Materials).In contrast, the same STB stimuli delivered at 65 dB SPL do not evokeVEMPs or activate the cortex [51,53]. Caloric and galvanic vestibularstimuli have been used in other fMRI studies of vestibular brain activi-ty [31], although their thermal and electrical sensations create undesir-able effects that are more difficult to control than non-vestibularresponses to STBs. For example, caloric and galvanic stimuli aremore likely to elicit head movements that degrade fMRI data.Hence, to investigate how personality traits influenced reactivity ofthe vestibular and anxiety systems, we compared responses to identicalSTBs at 100 dB and 65 dB SPL. To control for non-specific reactionsto sound, we also obtained brain responses to white-noise stimuli at100 dB SPL.
STBs had a frequency of 500 Hz, rise and fall times of 1 ms, pla-teau times of 8 ms, and were presented at repetition rates of 3 Hz,as recommended in a number of previous studies [59–61]. White-noise stimuli were sinusoidally modulated signals that reproducedthe time variability of STBs. In particular, white-noise had a loudnessthat varied at a frequency of 3 Hz. This prevented habituation towhite-noise stimuli while obtaining data to control for startle andother non-specific responses.
The experimental task included four stimuli: (1) STB100dB SPL;(2) STB65dB SPL; (3) white noise 100dB SPL; and (4) rest periodswith no stimulus presentation. Forty-five stimuli per typewere groupedin blocks lasting 15 sec each (rest blocks also lasted 15 sec). Foursessions (2 for the left and 2 for the right ear) including 4 blocks pertype of stimuli were presented in about 16 minutes. The order ofsessions and side of first stimulated ear were counterbalanced acrossall participants. To attenuate interferences from MRI scanner noise, weisolated the headphones with soundproof foam cushions and usedfoam plugs in the non-stimulated ear. Participants were asked to fixatea central point, attend to auditory stimuli, and report howmany types ofstimuli they heard. Following data acquisition, participants were askedto report the unpleasantness of each type of stimulus on an annoyancescale from 1 to 10 and any sensations of illusory body motion. All
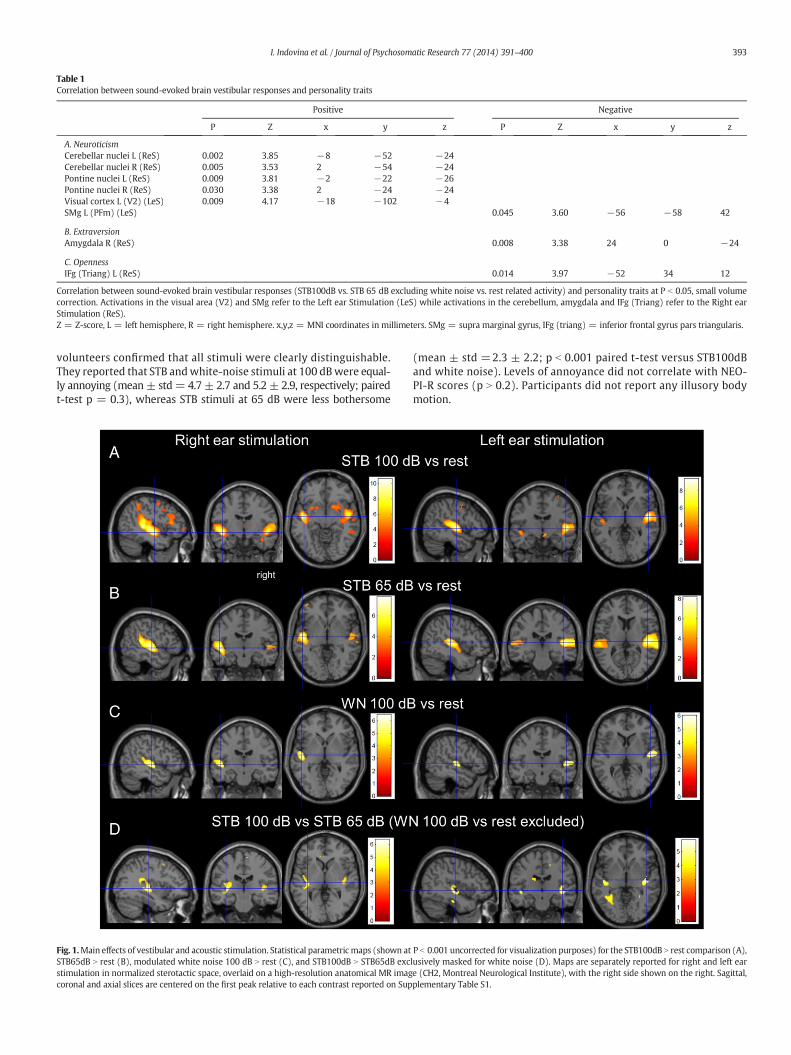
Table 1Correlation between sound-evoked brain vestibular responses and personality traits
Positive Negative
P Z x y z P Z x y z
A. NeuroticismCerebellar nuclei L (ReS) 0.002 3.85 −8 −52 −24Cerebellar nuclei R (ReS) 0.005 3.53 2 −54 −24Pontine nuclei L (ReS) 0.009 3.81 −2 −22 −26Pontine nuclei R (ReS) 0.030 3.38 2 −24 −24Visual cortex L (V2) (LeS) 0.009 4.17 −18 −102 −4SMg L (PFm) (LeS) 0.045 3.60 −56 −58 42
B. ExtraversionAmygdala R (ReS) 0.008 3.38 24 0 −24
C. OpennessIFg (Triang) L (ReS) 0.014 3.97 −52 34 12
Correlation between sound-evoked brain vestibular responses (STB100dB vs. STB 65 dB excluding white noise vs. rest related activity) and personality traits at P b 0.05, small volumecorrection. Activations in the visual area (V2) and SMg refer to the Left ear Stimulation (LeS) while activations in the cerebellum, amygdala and IFg (Triang) refer to the Right earStimulation (ReS).Z = Z-score, L = left hemisphere, R = right hemisphere. x,y,z = MNI coordinates in millimeters. SMg = supra marginal gyrus, IFg (triang) = inferior frontal gyrus pars triangularis.
393I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
volunteers confirmed that all stimuli were clearly distinguishable.They reported that STB andwhite-noise stimuli at 100 dBwere equal-ly annoying (mean ± std= 4.7 ± 2.7 and 5.2 ± 2.9, respectively; pairedt-test p = 0.3), whereas STB stimuli at 65 dB were less bothersome
Fig. 1.Main effects of vestibular and acoustic stimulation. Statistical parametricmaps (shown atSTB65dB N rest (B), modulated white noise 100 dB N rest (C), and STB100dB N STB65dB excstimulation in normalized sterotactic space, overlaid on a high-resolution anatomical MR imagcoronal and axial slices are centered on the first peak relative to each contrast reported on Sup
(mean ± std =2.3 ± 2.2; p b 0.001 paired t-test versus STB100dBand white noise). Levels of annoyance did not correlate with NEO-PI-R scores (p N 0.2). Participants did not report any illusory bodymotion.
P b 0.001 uncorrected for visualization purposes) for the STB100dB N rest comparison (A),lusively masked for white noise (D). Maps are separately reported for right and left eare (CH2, Montreal Neurological Institute), with the right side shown on the right. Sagittal,plementary Table S1.
394 I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
Image acquisition
fMRI was performed on a 3-T unit with an eight-channel headcoil. Head movements were minimized using foam pads aroundparticipants' head. No participant had head movements N2 mm.Whole-brain fMRI data were acquired with echo planar images sen-sitive to blood oxygenation level-dependent (BOLD) contrast (39axial slices, 3-mm thickness each; repetition time = 2000 ms;echo time = 30 ms; voxel size: 3×3×3 mm).
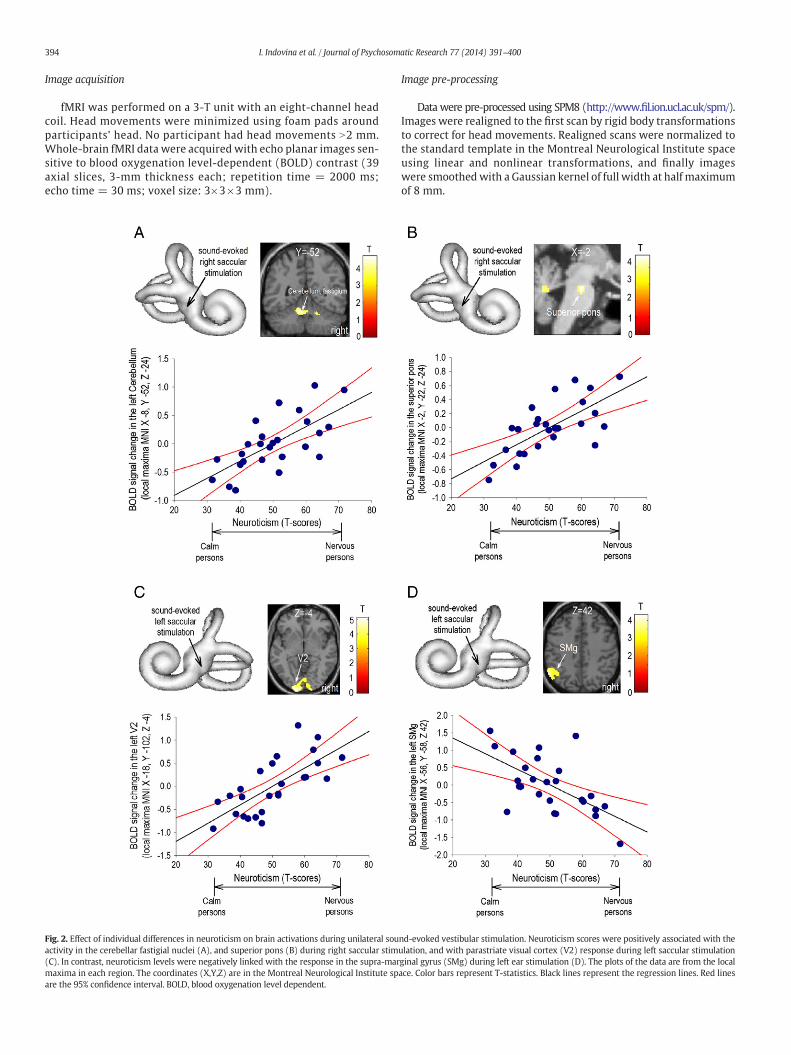
Fig. 2. Effect of individual differences in neuroticism on brain activations during unilateral souactivity in the cerebellar fastigial nuclei (A), and superior pons (B) during right saccular stim(C). In contrast, neuroticism levels were negatively linked with the response in the supra-marmaxima in each region. The coordinates (X,Y,Z) are in the Montreal Neurological Institute spare the 95% confidence interval. BOLD, blood oxygenation level dependent.
Image pre-processing
Data were pre-processed using SPM8 (http://www.fil.ion.ucl.ac.uk/spm/).Images were realigned to the first scan by rigid body transformationsto correct for head movements. Realigned scans were normalized tothe standard template in the Montreal Neurological Institute spaceusing linear and nonlinear transformations, and finally imageswere smoothedwith a Gaussian kernel of full width at half maximumof 8 mm.
nd-evoked vestibular stimulation. Neuroticism scores were positively associated with theulation, and with parastriate visual cortex (V2) response during left saccular stimulationginal gyrus (SMg) during left ear stimulation (D). The plots of the data are from the localace. Color bars represent T-statistics. Black lines represent the regression lines. Red lines
395I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
fMRI analysis of regional responses
For each participant, general linear models (GLMs) assessed region-ally specific effects of task parameters on BOLD activations. First-levelGLMs included 4 experimental conditions (STB100dB, STB65dB, white-noise 100dB, and rest) modeled as epochs of fixed duration and con-volved with the hemodynamic response function, and 6 realignmentparameters as effects of no interest to remove residual motion-relatedvariance. Low-frequency signal drift was eliminated using a high-passfilter (cut-off, 128 sec). Autoregressive AR(1) models corrected forvoxels' autocorrelations.
Subjects' specific contrast images (i.e., STB100dB N rest, STB65dB N
rest, white-noise 100dB N rest, STB100dB N STB65dB) were generatedseparately for right and left ears, averaging the two sessions of eachside, and entered into second-level GLMs investigating the main effectof the task per each contrast (one sample t-tests). Because white-noise stimuli were qualitatively different from STBs, they were not in-cluded in the comparison assessing brain activations associated withvestibular stimulation (i.e., the STB100dB N STB65dB contrast). Instead,white noise 100dB N rest contrast was used to exclusively mask theSTB100dB N STB65dB comparison. This way, we eliminated residual au-ditory activity that may have been present in the STB100dB N STB65dBcontrast as a result of differences in stimulus loudness. To create theexclusive mask from the white noise 100dB N rest contrast, we used alenient threshold of P ≤ 0.05, uncorrected. This threshold was chosen
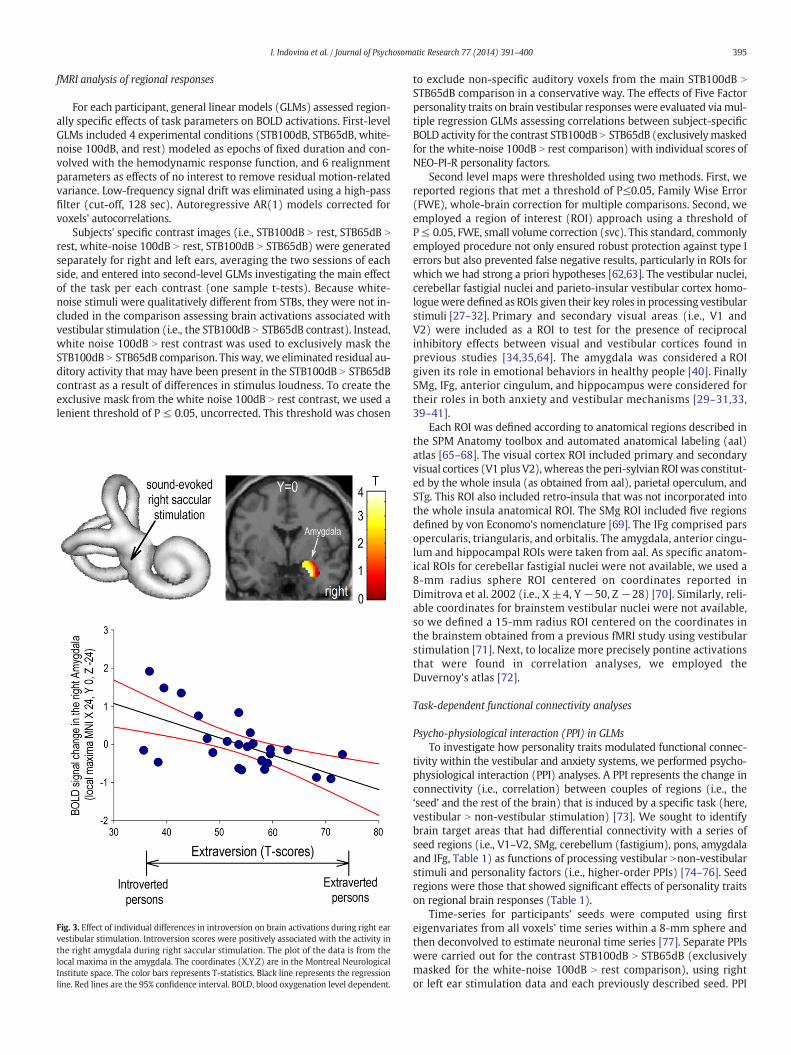
Fig. 3. Effect of individual differences in introversion on brain activations during right earvestibular stimulation. Introversion scores were positively associated with the activity inthe right amygdala during right saccular stimulation. The plot of the data is from thelocal maxima in the amygdala. The coordinates (X,Y,Z) are in the Montreal NeurologicalInstitute space. The color bars represents T-statistics. Black line represents the regressionline. Red lines are the 95% confidence interval. BOLD, blood oxygenation level dependent.
to exclude non-specific auditory voxels from the main STB100dB N
STB65dB comparison in a conservative way. The effects of Five Factorpersonality traits on brain vestibular responses were evaluated viamul-tiple regression GLMs assessing correlations between subject-specificBOLD activity for the contrast STB100dB N STB65dB (exclusivelymaskedfor the white-noise 100dB N rest comparison) with individual scores ofNEO-PI-R personality factors.
Second level maps were thresholded using two methods. First, wereported regions that met a threshold of P≤0.05, Family Wise Error(FWE), whole-brain correction for multiple comparisons. Second, weemployed a region of interest (ROI) approach using a threshold ofP≤ 0.05, FWE, small volume correction (svc). This standard, commonlyemployed procedure not only ensured robust protection against type Ierrors but also prevented false negative results, particularly in ROIs forwhich we had strong a priori hypotheses [62,63]. The vestibular nuclei,cerebellar fastigial nuclei and parieto-insular vestibular cortex homo-loguewere defined as ROIs given their key roles in processing vestibularstimuli [27–32]. Primary and secondary visual areas (i.e., V1 andV2) were included as a ROI to test for the presence of reciprocalinhibitory effects between visual and vestibular cortices found inprevious studies [34,35,64]. The amygdala was considered a ROIgiven its role in emotional behaviors in healthy people [40]. FinallySMg, IFg, anterior cingulum, and hippocampus were considered fortheir roles in both anxiety and vestibular mechanisms [29–31,33,39–41].
Each ROI was defined according to anatomical regions described inthe SPM Anatomy toolbox and automated anatomical labeling (aal)atlas [65–68]. The visual cortex ROI included primary and secondaryvisual cortices (V1 plus V2),whereas the peri-sylvian ROIwas constitut-ed by the whole insula (as obtained from aal), parietal operculum, andSTg. This ROI also included retro-insula that was not incorporated intothe whole insula anatomical ROI. The SMg ROI included five regionsdefined by von Economo's nomenclature [69]. The IFg comprised parsopercularis, triangularis, and orbitalis. The amygdala, anterior cingu-lum and hippocampal ROIs were taken from aal. As specific anatom-ical ROIs for cerebellar fastigial nuclei were not available, we used a8-mm radius sphere ROI centered on coordinates reported inDimitrova et al. 2002 (i.e., X ±4, Y −50, Z −28) [70]. Similarly, reli-able coordinates for brainstem vestibular nuclei were not available,so we defined a 15-mm radius ROI centered on the coordinates inthe brainstem obtained from a previous fMRI study using vestibularstimulation [71]. Next, to localize more precisely pontine activationsthat were found in correlation analyses, we employed theDuvernoy’s atlas [72].
Task-dependent functional connectivity analyses
Psycho-physiological interaction (PPI) in GLMsTo investigate how personality traits modulated functional connec-
tivity within the vestibular and anxiety systems, we performed psycho-physiological interaction (PPI) analyses. A PPI represents the change inconnectivity (i.e., correlation) between couples of regions (i.e., the‘seed’ and the rest of the brain) that is induced by a specific task (here,vestibular N non-vestibular stimulation) [73]. We sought to identifybrain target areas that had differential connectivity with a series ofseed regions (i.e., V1–V2, SMg, cerebellum (fastigium), pons, amygdalaand IFg, Table 1) as functions of processing vestibular Nnon-vestibularstimuli and personality factors (i.e., higher-order PPIs) [74–76]. Seedregions were those that showed significant effects of personality traitson regional brain responses (Table 1).
Time-series for participants' seeds were computed using firsteigenvariates from all voxels' time series within a 8-mm sphere andthen deconvolved to estimate neuronal time series [77]. Separate PPIswere carried out for the contrast STB100dB N STB65dB (exclusivelymasked for the white-noise 100dB N rest comparison), using rightor left ear stimulation data and each previously described seed. PPI
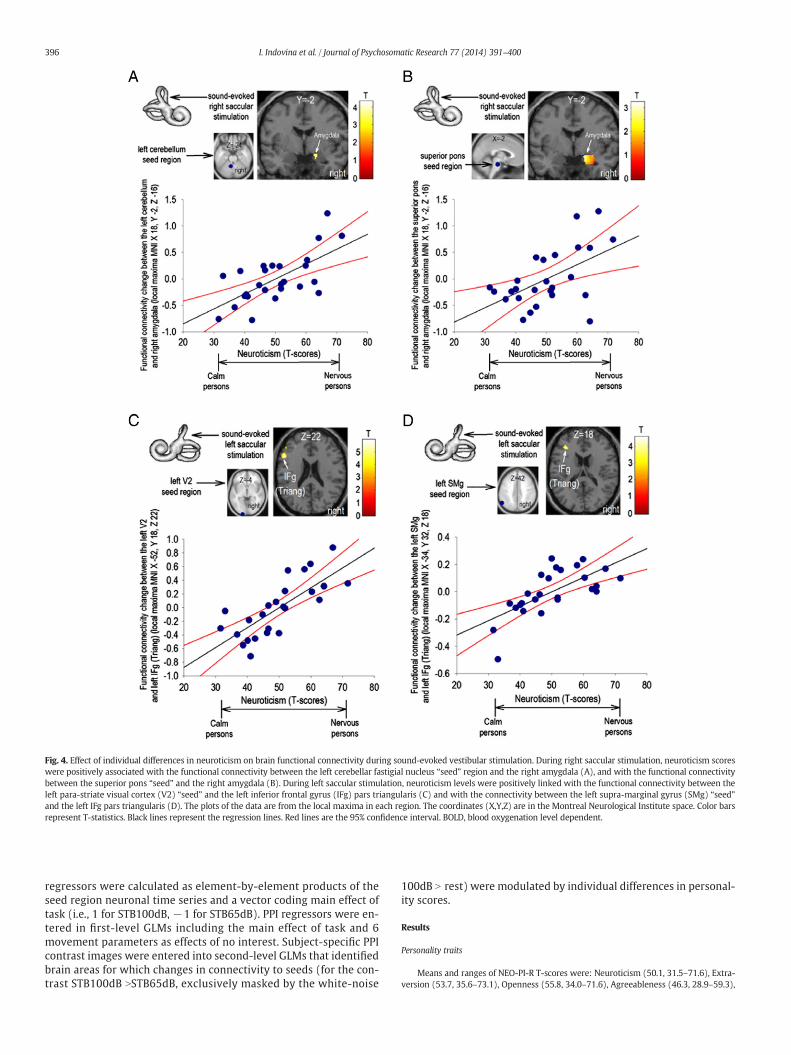
Fig. 4. Effect of individual differences in neuroticism on brain functional connectivity during sound-evoked vestibular stimulation. During right saccular stimulation, neuroticism scoreswere positively associated with the functional connectivity between the left cerebellar fastigial nucleus “seed” region and the right amygdala (A), and with the functional connectivitybetween the superior pons “seed” and the right amygdala (B). During left saccular stimulation, neuroticism levels were positively linked with the functional connectivity between theleft para-striate visual cortex (V2) “seed” and the left inferior frontal gyrus (IFg) pars triangularis (C) and with the connectivity between the left supra-marginal gyrus (SMg) “seed”and the left IFg pars triangularis (D). The plots of the data are from the local maxima in each region. The coordinates (X,Y,Z) are in the Montreal Neurological Institute space. Color barsrepresent T-statistics. Black lines represent the regression lines. Red lines are the 95% confidence interval. BOLD, blood oxygenation level dependent.
396 I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
regressors were calculated as element-by-element products of theseed region neuronal time series and a vector coding main effect oftask (i.e., 1 for STB100dB, −1 for STB65dB). PPI regressors were en-tered in first-level GLMs including the main effect of task and 6movement parameters as effects of no interest. Subject-specific PPIcontrast images were entered into second-level GLMs that identifiedbrain areas for which changes in connectivity to seeds (for the con-trast STB100dB NSTB65dB, exclusively masked by the white-noise
100dB N rest) were modulated by individual differences in personal-ity scores.
Results
Personality traits
Means and ranges of NEO-PI-R T-scores were: Neuroticism (50.1, 31.5–71.6), Extra-version (53.7, 35.6–73.1), Openness (55.8, 34.0–71.6), Agreeableness (46.3, 28.9–59.3),
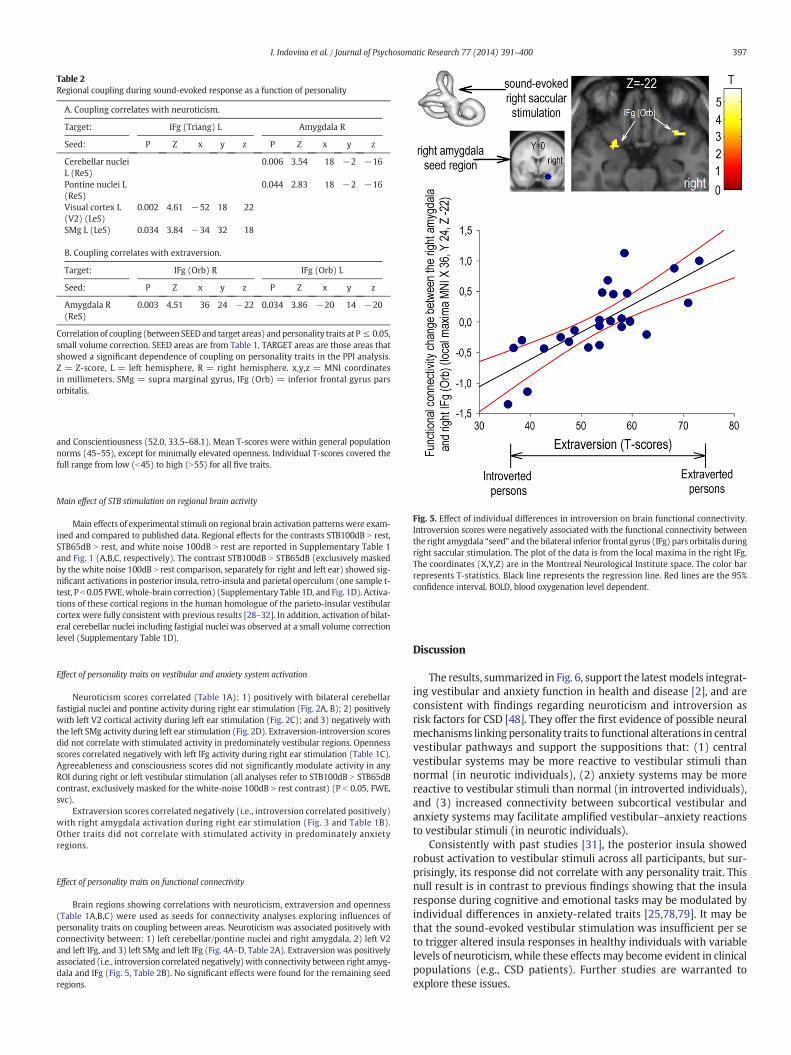
Table 2Regional coupling during sound-evoked response as a function of personality
A. Coupling correlates with neuroticism.
Target: IFg (Triang) L Amygdala R
Seed: P Z x y z P Z x y z
Cerebellar nucleiL (ReS)
0.006 3.54 18 −2 −16
Pontine nuclei L(ReS)
0.044 2.83 18 −2 −16
Visual cortex L(V2) (LeS)
0.002 4.61 −52 18 22
SMg L (LeS) 0.034 3.84 −34 32 18
B. Coupling correlates with extraversion.
Target: IFg (Orb) R IFg (Orb) L
Seed: P Z x y z P Z x y z
Amygdala R(ReS)
0.003 4.51 36 24 −22 0.034 3.86 −20 14 −20
Correlation of coupling (between SEEDand target areas) andpersonality traits at P ≤ 0.05,small volume correction. SEED areas are from Table 1, TARGET areas are those areas thatshowed a significant dependence of coupling on personality traits in the PPI analysis.Z = Z-score, L = left hemisphere, R = right hemisphere. x,y,z = MNI coordinatesin millimeters. SMg = supra marginal gyrus, IFg (Orb) = inferior frontal gyrus parsorbitalis.
397I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
and Conscientiousness (52.0, 33.5–68.1). Mean T-scores were within general populationnorms (45–55), except for minimally elevated openness. Individual T-scores covered thefull range from low (b45) to high (N55) for all five traits.
Fig. 5. Effect of individual differences in introversion on brain functional connectivity.Introversion scores were negatively associated with the functional connectivity betweenthe right amygdala “seed” and the bilateral inferior frontal gyrus (IFg) pars orbitalis duringright saccular stimulation. The plot of the data is from the local maxima in the right IFg.The coordinates (X,Y,Z) are in the Montreal Neurological Institute space. The color barrepresents T-statistics. Black line represents the regression line. Red lines are the 95%confidence interval. BOLD, blood oxygenation level dependent.
Main effect of STB stimulation on regional brain activity
Main effects of experimental stimuli on regional brain activation patternswere exam-ined and compared to published data. Regional effects for the contrasts STB100dB N rest,STB65dB N rest, and white noise 100dB N rest are reported in Supplementary Table 1and Fig. 1 (A,B,C, respectively). The contrast STB100dB N STB65dB (exclusively maskedby the white noise 100dB N rest comparison, separately for right and left ear) showed sig-nificant activations in posterior insula, retro-insula and parietal operculum (one sample t-test, P b 0.05 FWE,whole-brain correction) (Supplementary Table 1D, and Fig. 1D). Activa-tions of these cortical regions in the human homologue of the parieto-insular vestibularcortex were fully consistent with previous results [28–32]. In addition, activation of bilat-eral cerebellar nuclei including fastigial nuclei was observed at a small volume correctionlevel (Supplementary Table 1D).
Effect of personality traits on vestibular and anxiety system activation
Neuroticism scores correlated (Table 1A): 1) positively with bilateral cerebellarfastigial nuclei and pontine activity during right ear stimulation (Fig. 2A, B); 2) positivelywith left V2 cortical activity during left ear stimulation (Fig. 2C); and 3) negatively withthe left SMg activity during left ear stimulation (Fig. 2D). Extraversion-introversion scoresdid not correlate with stimulated activity in predominately vestibular regions. Opennessscores correlated negatively with left IFg activity during right ear stimulation (Table 1C).Agreeableness and consciousness scores did not significantly modulate activity in anyROI during right or left vestibular stimulation (all analyses refer to STB100dB N STB65dBcontrast, exclusively masked for the white-noise 100dB N rest contrast) (P b 0.05, FWE,svc).
Extraversion scores correlated negatively (i.e., introversion correlated positively)with right amygdala activation during right ear stimulation (Fig. 3 and Table 1B).Other traits did not correlate with stimulated activity in predominately anxietyregions.
Effect of personality traits on functional connectivity
Brain regions showing correlations with neuroticism, extraversion and openness(Table 1A,B,C) were used as seeds for connectivity analyses exploring influences ofpersonality traits on coupling between areas. Neuroticism was associated positively withconnectivity between: 1) left cerebellar/pontine nuclei and right amygdala, 2) left V2and left IFg, and 3) left SMg and left IFg (Fig. 4A–D, Table 2A). Extraversion was positivelyassociated (i.e., introversion correlated negatively)with connectivity between right amyg-dala and IFg (Fig. 5, Table 2B). No significant effects were found for the remaining seedregions.
Discussion
The results, summarized in Fig. 6, support the latestmodels integrat-ing vestibular and anxiety function in health and disease [2], and areconsistent with findings regarding neuroticism and introversion asrisk factors for CSD [48]. They offer the first evidence of possible neuralmechanisms linking personality traits to functional alterations in centralvestibular pathways and support the suppositions that: (1) centralvestibular systems may be more reactive to vestibular stimuli thannormal (in neurotic individuals), (2) anxiety systems may be morereactive to vestibular stimuli than normal (in introverted individuals),and (3) increased connectivity between subcortical vestibular andanxiety systems may facilitate amplified vestibular–anxiety reactionsto vestibular stimuli (in neurotic individuals).
Consistently with past studies [31], the posterior insula showedrobust activation to vestibular stimuli across all participants, but sur-prisingly, its response did not correlate with any personality trait. Thisnull result is in contrast to previous findings showing that the insularesponse during cognitive and emotional tasks may be modulated byindividual differences in anxiety-related traits [25,78,79]. It may bethat the sound-evoked vestibular stimulation was insufficient per seto trigger altered insula responses in healthy individuals with variablelevels of neuroticism, while these effectsmay become evident in clinicalpopulations (e.g., CSD patients). Further studies are warranted toexplore these issues.
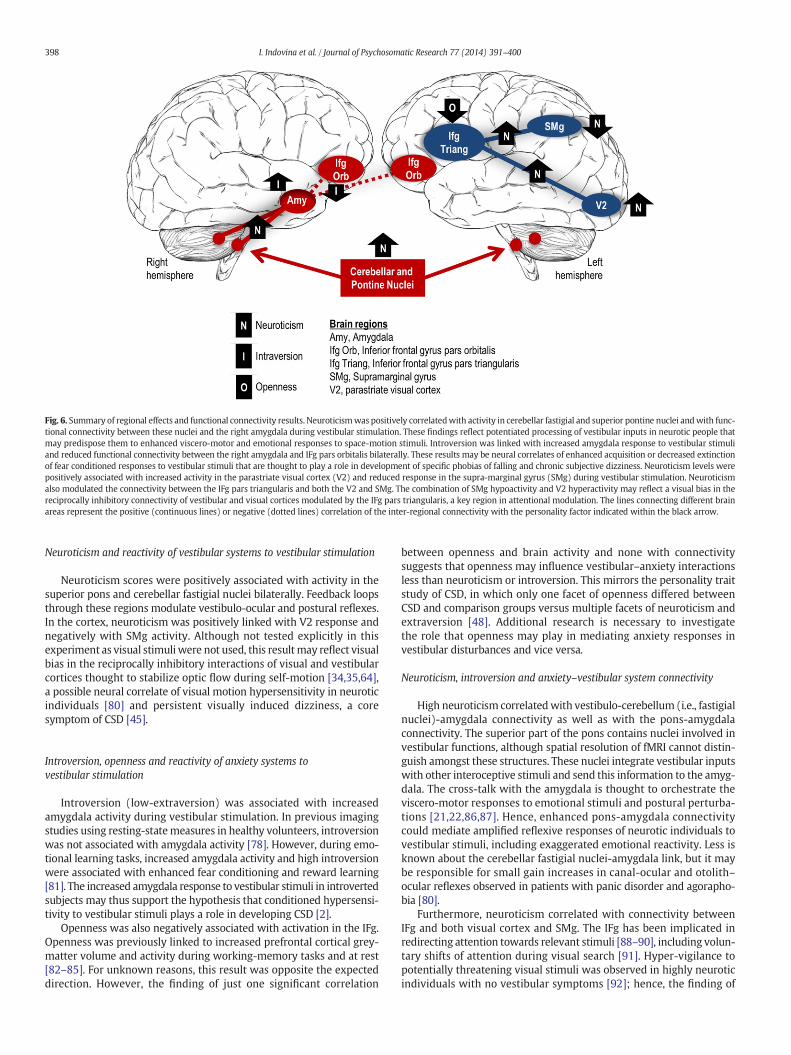
Fig. 6. Summary of regional effects and functional connectivity results. Neuroticismwas positively correlatedwith activity in cerebellar fastigial and superior pontine nuclei andwith func-tional connectivity between these nuclei and the right amygdala during vestibular stimulation. These findings reflect potentiated processing of vestibular inputs in neurotic people thatmay predispose them to enhanced viscero-motor and emotional responses to space-motion stimuli. Introversion was linked with increased amygdala response to vestibular stimuliand reduced functional connectivity between the right amygdala and IFg pars orbitalis bilaterally. These results may be neural correlates of enhanced acquisition or decreased extinctionof fear conditioned responses to vestibular stimuli that are thought to play a role in development of specific phobias of falling and chronic subjective dizziness. Neuroticism levels werepositively associated with increased activity in the parastriate visual cortex (V2) and reduced response in the supra-marginal gyrus (SMg) during vestibular stimulation. Neuroticismalso modulated the connectivity between the IFg pars triangularis and both the V2 and SMg. The combination of SMg hypoactivity and V2 hyperactivity may reflect a visual bias in thereciprocally inhibitory connectivity of vestibular and visual cortices modulated by the IFg pars triangularis, a key region in attentional modulation. The lines connecting different brainareas represent the positive (continuous lines) or negative (dotted lines) correlation of the inter-regional connectivity with the personality factor indicated within the black arrow.
398 I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
Neuroticism and reactivity of vestibular systems to vestibular stimulation
Neuroticism scores were positively associated with activity in thesuperior pons and cerebellar fastigial nuclei bilaterally. Feedback loopsthrough these regions modulate vestibulo-ocular and postural reflexes.In the cortex, neuroticism was positively linked with V2 response andnegatively with SMg activity. Although not tested explicitly in thisexperiment as visual stimuli were not used, this resultmay reflect visualbias in the reciprocally inhibitory interactions of visual and vestibularcortices thought to stabilize optic flow during self-motion [34,35,64],a possible neural correlate of visual motion hypersensitivity in neuroticindividuals [80] and persistent visually induced dizziness, a coresymptom of CSD [45].
Introversion, openness and reactivity of anxiety systems tovestibular stimulation
Introversion (low-extraversion) was associated with increasedamygdala activity during vestibular stimulation. In previous imagingstudies using resting-statemeasures in healthy volunteers, introversionwas not associated with amygdala activity [78]. However, during emo-tional learning tasks, increased amygdala activity and high introversionwere associated with enhanced fear conditioning and reward learning[81]. The increased amygdala response to vestibular stimuli in introvertedsubjects may thus support the hypothesis that conditioned hypersensi-tivity to vestibular stimuli plays a role in developing CSD [2].
Openness was also negatively associated with activation in the IFg.Openness was previously linked to increased prefrontal cortical grey-matter volume and activity during working-memory tasks and at rest[82–85]. For unknown reasons, this result was opposite the expecteddirection. However, the finding of just one significant correlation
between openness and brain activity and none with connectivitysuggests that openness may influence vestibular–anxiety interactionsless than neuroticism or introversion. This mirrors the personality traitstudy of CSD, in which only one facet of openness differed betweenCSD and comparison groups versus multiple facets of neuroticism andextraversion [48]. Additional research is necessary to investigatethe role that openness may play in mediating anxiety responses investibular disturbances and vice versa.
Neuroticism, introversion and anxiety–vestibular system connectivity
High neuroticism correlatedwith vestibulo-cerebellum (i.e., fastigialnuclei)-amygdala connectivity as well as with the pons-amygdalaconnectivity. The superior part of the pons contains nuclei involved investibular functions, although spatial resolution of fMRI cannot distin-guish amongst these structures. These nuclei integrate vestibular inputswith other interoceptive stimuli and send this information to the amyg-dala. The cross-talk with the amygdala is thought to orchestrate theviscero-motor responses to emotional stimuli and postural perturba-tions [21,22,86,87]. Hence, enhanced pons-amygdala connectivitycould mediate amplified reflexive responses of neurotic individuals tovestibular stimuli, including exaggerated emotional reactivity. Less isknown about the cerebellar fastigial nuclei-amygdala link, but it maybe responsible for small gain increases in canal-ocular and otolith–ocular reflexes observed in patients with panic disorder and agorapho-bia [80].
Furthermore, neuroticism correlated with connectivity betweenIFg and both visual cortex and SMg. The IFg has been implicated inredirecting attention towards relevant stimuli [88–90], including volun-tary shifts of attention during visual search [91]. Hyper-vigilance topotentially threatening visual stimuli was observed in highly neuroticindividuals with no vestibular symptoms [92]; hence, the finding of

399I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
reduced SMg activity and increased V2 response to STB100dB stimuliraises the possibility of visual bias in multi-sensory integration ofspace-motion stimuli in neurotic people.
Introversion was negatively associated (extraversion positively cor-related) with amgydala-IFg (the orbitalis part) connectivity. Extensiveconnections between amygdala and IFg are important for extinguishingfear responses to conditioned stimuli [41,93–96]. Imaging studies foundreduced connectivity between amygdala and frontal cortical areas insocial and generalized anxiety disorders [78,81], suggesting that re-duced amygdala-prefrontal cortical connectivity may reflect a generalproneness to anxiety-related disorders in introverts.
Although CSD was previously independently linked to neuroticismand introversion, the two traits together separated CSD most stronglyfrom other neurotologic/anxiety disorders [48]. Therefore, the risk ofCSD may be conveyed by vestibular and anxiety systems that simulta-neously express heightened reactivity to vestibular stimulation,mediat-ed by neuroticism and introversion, respectively. Personality traits havenot been studied as explicitly in other anxiety-related vestibulardisorders, though the converse of neuroticism and introversion(i.e., resilience and sense of coherence) predicted fewer psychosomaticsequelae of acute vestibular syndromes [49].
This study provides the first evidence that individual differences inneuroticism, introversion, and possibly openness influence activityand functional connectivity within central vestibular and anxietysystems in a pattern consistent with published models of vestibular-anxiety interactions from brainstem to cortex [21,22,48]. Strengths ofthis investigation included use of a previously well-developed experi-mental paradigm to evoke vestibular responses, and a study samplewith a full spectrum of personality traits compatible with populationnorms, which provided valid data to test the personality modulationhypotheses. The main limitation is that any correlates between brainactivation patterns and clinical phenotypes must be interpreted conser-vatively until they can be replicated and extended. For personality traitmodulation of vestibular responses that will require testing of addition-al visual and vestibular stimuli in larger cohorts of normal individualsand patients with anxiety-related vestibular disorders (e.g., CSD).Nevertheless, the concordance of these results with existing humanand animal data and recent neuroanatomical models is promising.
Financial support
Italian Ministry of University and Research (PRIN grant), NationalResearch Council.
Financial disclosures
None of the authors have conflicts of interests.
Acknowledgments
We thank our volunteers for kindly participating in this study andDr. Martin Vestergaard for helping with auditory stimuli production.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.jpsychores.2014.09.005.
References
[1] Balaban CD, Jacob RG. Background and history of the interface between anxiety andvertigo. J Anxiety Disord 2001;15:27–51.
[2] Staab JP, Balaban CD, Furman JM. Threat assessment and locomotion: clinical appli-cations of an integrated model of anxiety and postural control. Semin Neurol 2013;33:297–306.
[3] Carpenter MG, Frank JS, Silcher CP. Surface height effects on postural control: ahypothesis for a stiffness strategy for stance. J Vestib Res 1999;9:277–86.
[4] Adkin AL, Frank JS, Carpenter MG, Peysar GW. Postural control is scaled to level ofpostural threat. Gait Posture 2000;12:87–93.
[5] Brown LA, Frank JS. Postural compensations to the potential consequences ofinstability: kinematics. Gait Posture 1997;6:89–97.
[6] Davis JR, Campbell AD, Adkin AL, Carpenter MG. The relationship between fear offalling and human postural control. Gait Posture 2009;29:275–9.
[7] Davis JR, Horslen BC, Nishikawa K, Fukushima K, Chua R, Inglis JT, et al. Humanproprioceptive adaptations during states of height-induced fear and anxiety. JNeurophysiol 2011;106:3082–90.
[8] Cleworth TW, Horslen BC, Carpenter MG. Influence of real and virtual heights onstanding balance. Gait Posture 2012;36:172–6.
[9] Adkin AL, Frank JS, Carpenter MG, Peysar GW. Fear of falling modifies anticipatorypostural control. Exp Brain Res 2002;143:160–70.
[10] Hainaut J-P, Caillet G, Lestienne FG, Bolmont B. The role of trait anxiety on staticbalance performance in control and anxiogenic situations. Gait Posture 2011;33:604–8.
[11] Eckhardt A, Tettenborn B, Krauthauser H, Thomalske C, Hartmann O, Hoffmann SO,et al. Vertigo and anxiety disorders – results of interdisciplinary evaluation.Laryngorhinootologie 1996;75:517–22.
[12] Eckhardt-Henn A, Breuer P, Thomalske C, Hoffmann SO, Hopf HC. Anxiety disordersand other psychiatric subgroups in patients complaining of dizziness. J AnxietyDisord 2003;17:369–88.
[13] Clark DB, Hirsch BE, Smith MG, Furman JM, Jacob RG. Panic in otolaryngology pa-tients presenting with dizziness or hearing loss. Am J Psychiatry 1994;151:1223–5.
[14] Persoons P, Luyckx K, Desloovere C, Vandenberghe J, Fischler B. Anxiety and mooddisorders in otorhinolaryngology outpatients presenting with dizziness: validationof the self-administered PRIME-MD Patient Health Questionnaire and epidemiology.Gen Hosp Psychiatry 2003;25:316–23.
[15] Eagger S, Luxon LM, Davies RA, Coelho A, Ron MA. Psychiatric morbidity in patientswith peripheral vestibular disorder: a clinical and neuro-otological study. J NeurolNeurosurg Psychiatry 1992;55:383–7.
[16] Kammerlind A-SC, Ledin TEA, Skargren EIB, Odkvist LM. Long-term follow-up afteracute unilateral vestibular loss and comparison between subjects with and withoutremaining symptoms. Acta Otolaryngol (Stockh) 2005;125:946–53.
[17] Celestino D, Rosini E, Carucci ML, Marconi PL, Vercillo E. Ménière's disease andanxiety disorders. Acta Otorhinolaryngol Ital 2003;23:421–7.
[18] Best C, Eckhardt-Henn A, Tschan R, Dieterich M. Psychiatric morbidity and comor-bidity in different vestibular vertigo syndromes. Results of a prospective longitudinalstudy over one year. J Neurol 2009;256:58–65.
[19] Boleas-Aguirre MS, Sánchez-Ferrandiz N, Guillén-Grima F, Perez N. Long-term dis-ability of class A patients with Ménière's disease after treatment with intratympanicgentamicin. Laryngoscope 2007;117:1474–81.
[20] Staab JP. Chronic dizziness: the interface between psychiatry and neuro-otology.Curr Opin Neurol 2006;19:41–8.
[21] Balaban CD. Neural substrates linking balance control and anxiety. Physiol Behav2002;77:469–75.
[22] Balaban CD. Projections from the parabrachial nucleus to the vestibular nuclei: po-tential substrates for autonomic and limbic influences on vestibular responses.Brain Res 2004;996:126–37.
[23] Balaban CD, Thayer JF. Neurological bases for balance-anxiety links. J Anxiety Disord2001;15:53–79.
[24] Gurvich C, Maller JJ, Lithgow B, Haghgooie S, Kulkarni J. Vestibular insights intocognition and psychiatry. Brain Res 2013;1537:244–59.
[25] Paulus MP, Stein MB. An insular view of anxiety. Biol Psychiatry 2006;60:383–7.[26] Viaud-Delmon I, Venault P, Chapouthier G. Behavioral models for anxiety and
multisensory integration in animals and humans. Prog NeuropsychopharmacolBiol Psychiatry 2011;35:1391–9.
[27] Angelaki DE, Cullen KE. Vestibular system: the many facets of a multimodal sense.Annu Rev Neurosci 2008;31:125–50.
[28] Brandt T, Dieterich M. The vestibular cortex. Its locations, functions, and disorders.Ann N Y Acad Sci 1999;871:293–312.
[29] Zu Eulenburg P, Caspers S, Roski C, Eickhoff SB. Meta-analytical definition andfunctional connectivity of the human vestibular cortex. Neuroimage 2012;60:162–9.
[30] Indovina I, Maffei V, Bosco G, Zago M, Macaluso E, Lacquaniti F. Representationof visual gravitational motion in the human vestibular cortex. Science 2005;308:416–9.
[31] Lopez C, Blanke O, Mast FW. The human vestibular cortex revealed by coordinate-based activation likelihood estimationmeta-analysis. Neuroscience 2012;212:159–79.
[32] Lopez C, Blanke O. The thalamocortical vestibular system in animals and humans.Brain Res Rev 2011;67:119–46.
[33] Hüfner K, Strupp M, Smith P, Brandt T, Jahn K. Spatial separation of visual andvestibular processing in the human hippocampal formation. Ann N Y Acad Sci2011;1233:177–86.
[34] Brandt T. Cortical visual–vestibular interaction for spatial orientation and self-motion perception. Curr Opin Neurol 1999;12:1–4.
[35] Brandt T, Bartenstein P, Janek A, Dieterich M. Reciprocal inhibitory visual–vestibularinteraction. Visual motion stimulation deactivates the parieto-insular vestibularcortex. Brain 1998;121:1749–58.
[36] Bremmer F, Schlack A, Shah NJ, Zafiris O, Kubischik M, Hoffmann K, et al. Polymodalmotion processing in posterior parietal and premotor cortex: a human fMRI studystrongly implies equivalencies between humans and monkeys. Neuron 2001;29:287–96.
[37] Britten KH. Mechanisms of self-motion perception. Annu Rev Neurosci 2008;31:389–410.
[38] De Waele C, Baudonnière PM, Lepecq JC, Tran Ba Huy P, Vidal PP. Vestibular projec-tions in the human cortex. Exp Brain Res 2001;141:541–51.

400 I. Indovina et al. / Journal of Psychosomatic Research 77 (2014) 391–400
[39] Sylvester CM, Corbetta M, Raichle ME, Rodebaugh TL, Schlaggar BL, Sheline YI, et al.Functional network dysfunction in anxiety and anxiety disorders. Trends Neurosci2012;35:527–35.
[40] Bishop SJ. Neurocognitive mechanisms of anxiety: an integrative account. TrendsCogn Sci 2007;11:307–16.
[41] Indovina I, Robbins TW, Núñez-Elizalde AO, Dunn BD, Bishop SJ. Fear-conditioningmechanisms associated with trait vulnerability to anxiety in humans. Neuron2011;69:563–71.
[42] Brandt T. Phobic postural vertigo. Neurology 1996;46:1515–9.[43] Brandt T, Dieterich M. Phobischer Attacken-Schwank-schwindel, ein neues
Syndrom? Munch Med Wochenschr 1986;28:247–50.[44] Staab JP, Ruckenstein MJ. Expanding the differential diagnosis of chronic dizziness.
Arch Otolaryngol Head Neck Surg 2007;133:170–6.[45] Staab JP. Chronic subjective dizziness. Continuum(MinneapMinn) 2012;18:1118–41.[46] Staab JP, Ruckenstein MJ. Which comes first? Psychogenic dizziness versus otogenic
anxiety. Laryngoscope 2003;113:1714–8.[47] Staab JP, Ruckenstein MJ. Chronic dizziness and anxiety: effect of course of illness on
treatment outcome. Arch Otolaryngol Head Neck Surg 2005;131:675–9.[48] Staab JP, Rohe DE, Eggers SDZ, Shepard NT. Anxious, introverted personality traits in
patients with chronic subjective dizziness. J Psychosom Res 2014;76:80–3.[49] Tschan R, Best C, Beutel ME, Knebel A, Wiltink J, Dieterich M, et al. Patients' psycho-
logical well-being and resilient coping protect from secondary somatoform vertigoand dizziness (SVD) 1 year after vestibular disease. J Neurol 2011;258:104–12.
[50] Costa Jr PT, McCrae RR. Stability and change in personality assessment: the revisedNEO Personality Inventory in the year 2000. J Pers Assess 1997;68:86–94.
[51] Janzen J, Schlindwein P, Bense S, Bauermann T, Vucurevic G, Stoeter P, et al. Neuralcorrelates of hemispheric dominance and ipsilaterality within the vestibular system.Neuroimage 2008;42:1508–18.
[52] Miyamoto T, Fukushima K, Takada T, de Waele C, Vidal P-P. Saccular stimulation ofthe human cortex: a functional magnetic resonance imaging study. Neurosci Lett2007;423:68–72.
[53] Schlindwein P, Mueller M, Bauermann T, Brandt T, Stoeter P, Dieterich M. Corticalrepresentation of saccular vestibular stimulation: VEMPs in fMRI. Neuroimage2008;39:19–31.
[54] Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, et al. The Mini-International Neuropsychiatric Interview (M.I.N.I.): the development and validationof a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychi-atry 1998;59:22–33 [quiz 34–57].
[55] Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory.Neuropsychologia 1971;9:97–113.
[56] Brantberg K. Vestibular evoked myogenic potentials (VEMPs): usefulness in clinicalneurotology. Semin Neurol 2009;29:541–7.
[57] Colebatch JG, Halmagyi GM, Skuse NF. Myogenic potentials generated by a click-evoked vestibulocollic reflex. J Neurol Neurosurg Psychiatry 1994;57:190–7.
[58] Isaradisaikul S, Navacharoen N, Hanprasertpong C, Kangsanarak J. Cervicalvestibular-evoked myogenic potentials: norms and protocols. Int J Otolaryngol2012;2012:913515.
[59] Akin FW, Murnane OD, Proffitt TM. The effects of click and tone-burst stimulusparameters on the vestibular evoked myogenic potential (VEMP). J Am Acad Audiol2003;14:500–9 [quiz 534–535].
[60] Cheng PW, Murofushi T. The effect of rise/fall time on vestibular-evoked myogenicpotential triggered by short tone bursts. Acta Otolaryngol (Stockh) 2001;121:696–9.
[61] Cheng PW, Murofushi T. The effects of plateau time on vestibular-evoked myogenicpotentials triggered by tone bursts. Acta Otolaryngol (Stockh) 2001;121:935–8.
[62] Friston KJ. Testing for anatomically specified regional effects. HumBrainMapp 1997;5:133–6.
[63] Worsley KJ, Marrett S, Neelin P, Vandal AC, Friston KJ, Evans AC. A unified statisticalapproach for determining significant signals in images of cerebral activation. HumBrain Mapp 1996;4:58–73.
[64] Bense S, Stephan T, Yousry TA, Brandt T, Dieterich M. Multisensory cortical signalincreases anddecreases during vestibular galvanic stimulation (fMRI). J Neurophysiol2001;85:886–99.
[65] Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, Delcroix N,et al. Automated anatomical labeling of activations in SPM using a macroscopicanatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002;15:273–89.
[66] Eickhoff SB, Stephan KE, Mohlberg H, Grefkes C, Fink GR, Amunts K, et al. A new SPMtoolbox for combining probabilistic cytoarchitectonic maps and functional imagingdata. Neuroimage 2005;25:1325–35.
[67] Eickhoff SB, Heim S, Zilles K, Amunts K. Testing anatomically specified hypotheses infunctional imaging using cytoarchitectonic maps. Neuroimage 2006;32:570–82.
[68] Eickhoff SB, Paus T, Caspers S, Grosbras M-H, Evans AC, Zilles K, et al. Assignment offunctional activations to probabilistic cytoarchitectonic areas revisited. Neuroimage2007;36:511–21.
[69] von Economo C, Koskinas GN. Atlas of cytoarchitectonics of the adult humancerebral cortex. In: Triarhou LC, editor. Basel: Karger; 2008 [trans, rev].
[70] Dimitrova A, Weber J, Redies C, Kindsvater K, Maschke M, Kolb FP, et al. MRI atlas ofthe human cerebellar nuclei. Neuroimage 2002;17:240–55.
[71] Stephan T, Deutschländer A, Nolte A, Schneider E, Wiesmann M, Brandt T, et al.Functional MRI of galvanic vestibular stimulation with alternating currents at differ-ent frequencies. Neuroimage 2005;26:721–32.
[72] Naidich TP, Duvernoy HM, Delman BN, Sorensen AG, Kollias SS, Haacke EM.Duvernoy's Atlas of the Human Brain Stem and Cerebellum. Springer; 2009.
[73] Friston KJ, Buechel C, Fink GR, Morris J, Rolls E, Dolan RJ. Psychophysiological andmodulatory interactions in neuroimaging. Neuroimage 1997;6:218–29.
[74] Passamonti L, Rowe JB, Ewbank M, Hampshire A, Keane J, Calder AJ. Connectivityfrom the ventral anterior cingulate to the amygdala is modulated by appetitivemotivation in response to facial signals of aggression. Neuroimage 2008;43:562–70.
[75] Passamonti L, Rowe JB, Schwarzbauer C, Ewbank MP, von dem Hagen E, Calder AJ.Personality predicts the brain's response to viewing appetizing foods: the neuralbasis of a risk factor for overeating. J Neurosci 2009;29:43–51.
[76] Passamonti L, Crockett MJ, Apergis-Schoute AM, Clark L, Rowe JB, Calder AJ, et al.Effects of Acute Tryptophan Depletion on Prefrontal-Amygdala Connectivity WhileViewing Facial Signals of Aggression. Biol Psychiatry 2012;71:36–43.
[77] Gitelman DR, Penny WD, Ashburner J, Friston KJ. Modeling regional and psycho-physiologic interactions in fMRI: the importance of hemodynamic deconvolution.Neuroimage 2003;19:200–7.
[78] Deckersbach T, Miller KK, Klibanski A, Fischman A, Dougherty DD, Blais MA, et al.Regional cerebral brain metabolism correlates of neuroticism and extraversion.Depress Anxiety 2006;23:133–8.
[79] Paulus MP, Rogalsky C, Simmons A, Feinstein JS, Stein MB. Increased activation in theright insula during risk-taking decision making is related to harm avoidance andneuroticism. Neuroimage 2003;19:1439–48.
[80] Jacob RG, Redfern MS, Furman JM. Space and motion discomfort and abnormalbalance control in patients with anxiety disorders. J Neurol Neurosurg Psychiatry2009;80:74–8.
[81] Hooker CI, Verosky SC, Miyakawa A, Knight RT, D'Esposito M. The influence ofpersonality on neural mechanisms of observational fear and reward learning.Neuropsychologia 2008;46:2709–24.
[82] Sutin AR, Beason-Held LL, Resnick SM, Costa PT. Sex differences in resting-stateneural correlates of openness to experience among older adults. Cereb Cortex2009;19:2797–802.
[83] DeYoung CG, Shamosh NA, Green AE, Braver TS, Gray JR. Intellect as distinct fromOpenness: differences revealed by fMRI of working memory. J Pers Soc Psychol2009;97:883–92.
[84] Kapogiannis D, Sutin A, Davatzikos C, Costa Jr P, Resnick S. The five factors of person-ality and regional cortical variability in the Baltimore longitudinal study of aging.Hum Brain Mapp 2013;34:2829–40.
[85] Hu X, Erb M, Ackermann H, Martin JA, Grodd W, Reiterer SM. Voxel-basedmorphometry studies of personality: issue of statistical model specification–effectof nuisance covariates. Neuroimage 2011;54:1994–2005.
[86] Halberstadt AL, Balaban CD. Anterograde tracing of projections from the dorsalraphe nucleus to the vestibular nuclei. Neuroscience 2006;143:641–54.
[87] Halberstadt AL, Balaban CD. Selective anterograde tracing of the individual seroto-nergic and nonserotonergic components of the dorsal raphe nucleus projection tothe vestibular nuclei. Neuroscience 2007;147:207–23.
[88] Corbetta M, Patel G, Shulman GL. The reorienting system of the human brain: fromenvironment to theory of mind. Neuron 2008;58:306–24.
[89] Indovina I, Macaluso E. Occipital-parietal interactions during shifts of exogenousvisuospatial attention: trial-dependent changes of effective connectivity. MagnReson Imaging 2004;22:1477–86.
[90] Indovina I, Macaluso E. Dissociation of stimulus relevance and saliency factorsduring shifts of visuospatial attention. Cereb Cortex 2007;17:1701–11.
[91] Fairhall SL, Indovina I, Driver J, Macaluso E. The brain network underlying serialvisual search: comparing overt and covert spatial orienting, for activations and foreffective connectivity. Cereb Cortex 2009;19:2946–58.
[92] Staab JP. The influence of anxiety on ocular motor control and gaze. Curr OpinNeurol 2014;27:118–24.
[93] Delgado MR, Nearing KI, Ledoux JE, Phelps EA. Neural circuitry underlying the regu-lation of conditioned fear and its relation to extinction. Neuron 2008;59:829–38.
[94] Hartley CA, Phelps EA. Changing fear: the neurocircuitry of emotion regulation.Neuropsychopharmacology 2010;35:136–46.
[95] Milad MR, Wright CI, Orr SP, Pitman RK, Quirk GJ, Rauch SL. Recall of fear extinctionin humans activates the ventromedial prefrontal cortex and hippocampus inconcert. Biol Psychiatry 2007;62:446–54.
[96] Ochsner KN, Gross JJ. The cognitive control of emotion. Trends Cogn Sci 2005;9:242–9.
Related Documents











