Molecular Microbiology (2004) 52(2), 515–527 doi:10.1111/j.1365-2958.2004.03992.x © 2004 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2004 ? 2004522515527Original ArticleIsoprenoid lipid biosynthesis in archaeaY. Boucher, M. Kamekura and W. F. Doolittle Accepted 22 December, 2003. *For correspondence. E-mail [email protected]; Tel. (+1) 902 494 2968; Fax (+1) 902 494 1355. Origins and evolution of isoprenoid lipid biosynthesis in archaea Yan Boucher, 1 * Masahiro Kamekura 2 and W. Ford Doolittle 1 1 Canadian Institute for Advanced Research, Dalhousie University, Halifax, Nova Scotia, 5859 University Avenue, B3H 4H7 Canada. 2 Noda Institute for Scientific Research, 399 Noda. Noda- shi, Chiba-ken 278–0037 Japan. Summary A characteristic feature of the domain archaea are the lipids forming the hydrophobic core of their cell mem- brane. These unique lipids are composed of iso- prenoid side-chains stereospecifically ether linked to sn -glycerol-1-phosphate. Recently, considerable progress has been made in characterizing the enzymes responsible for the synthesis of archaeal lipids. However, little is known about their evolution. To better understand how this unique biosynthetic apparatus came to be, large-scale database surveys and phylogenetic analyses were performed. All char- acterized enzymes involved in the biosynthesis of isoprenoid side-chains and the glycerol phosphate backbone along with their assembly in ether lipids were included in these analyses. The sequence data available in public databases was complemented by an in-depth sampling of isoprenoid lipid biosynthesis genes from multiple genera of the archaeal order Halobacteriales, allowing us to look at the evolution of these enzymes on a smaller phylogenetic scale. This investigation of the isoprenoid biosynthesis apparatus of archaea on small and large phylogenetic scales reveals that it evolved through a combination of evolutionary processes, including the co-option of ancestral enzymes, modification of enzymatic specificity, orthologous and non-orthologous gene displacement, integration of components from eukaryotes and bacteria and lateral gene transfer within and between archaeal orders. Introduction Archaeal membrane lipids have several interesting char- acteristics which distinguish them from their bacterial and eukaryotic counterparts: (i) isoprenoid, not fatty acids, side-chains; (ii) ether, not ester, links joining these side- chains to the glycerol phosphate backbone; and (iii) the sn -1, not sn -3, stereochemistry of this backbone. Archaeal lipids side-chains, like all other isoprenoids, are assembled from two universal precursors: isopentenyl diphosphate (IPP) and its isomer dimethylallyl diphos- phate (DMAPP). Most eukaryotes and some bacteria syn- thesize these precursors through the mevalonate pathway (Boucher and Doolittle, 2000; Lange et al ., 2000). This pathway has been described in details in these organisms and is always composed of five steps, as illustrated in Fig. 1: (i) conversion of acetyl-CoA and acetoacetyl-CoA to 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA); (ii) reduc- tion of HMG-CoA to mevalonate; (iii) phosphorylation of mevalonate; (iv) phosphorylation of phosphomevalonate; and (v) conversion of diphosphomevalonate to IPP. Archaea also derive their isoprenoids from mevalonate, as shown by tracer studies more than two decades ago (Kates and Kushwaha, 1978). Orthologues of the enzymes catalysing the first three steps of the pathway in bacteria and eukaryotes can also be found in archaea (Smit and Mushegian, 2000). However, the last two enzymes of the standard bacterial/eukaryotic mevalonate pathway (phosphomevalonate kinase or PMK and diphos- phomevalonate decarboxylase or PPMD) are missing in most archaea. Only one exception to this is known, which is the identification of PPMD gene in the genome of the extremely halophilic archaeon Halobacterium sp. NRC-1 (Ng et al ., 2000). A type 1 isopentenyl diphosphate isomerase (IDI1), the enzyme responsible for the conver- sion of IPP to DMAPP in most eukaryotes and bacteria but generally absent from archaea, was also found in this genome. In some archaea, this enzymatic reaction could be catalysed by a functional analogue of IDI1, the non- homologous type 2 isopentenyl diphosphate isomerase (IDI2). Kaneda et al . (2001) have shown IDI2 to be part of the mevalonate pathway gene cluster of the actino- mycete Kitasatospora griseola and identified its presence in several other bacteria and archaea. The products of the mevalonate pathway (IPP and DMAPP) are used by archaea as building blocks for lipid side-chains. In acyclic lipids, these side-chains are com- posed of 20 or 25 carbons (C20 or C25). Such a length is reached through the sequential condensation of IPP

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Microbiology (2004)
52
(2), 515–527 doi:10.1111/j.1365-2958.2004.03992.x
© 2004 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd, 2004
? 2004
52
2515527
Original Article
Isoprenoid lipid biosynthesis in archaeaY. Boucher, M. Kamekura and W. F. Doolittle
Accepted 22 December, 2003. *For correspondence. [email protected]; Tel. (+1) 902 494 2968; Fax (+1) 902 494 1355.
Origins and evolution of isoprenoid lipid biosynthesis in archaea
Yan Boucher,
1
* Masahiro Kamekura
2
and W. Ford Doolittle
1
1
Canadian Institute for Advanced Research, Dalhousie University, Halifax, Nova Scotia, 5859 University Avenue, B3H 4H7 Canada.
2
Noda Institute for Scientific Research, 399 Noda. Noda-shi, Chiba-ken 278–0037 Japan.
Summary
A characteristic feature of the domain archaea are thelipids forming the hydrophobic core of their cell mem-brane. These unique lipids are composed of iso-prenoid side-chains stereospecifically ether linkedto
sn
-glycerol-1-phosphate. Recently, considerableprogress has been made in characterizing theenzymes responsible for the synthesis of archaeallipids. However, little is known about their evolution.To better understand how this unique biosyntheticapparatus came to be, large-scale database surveysand phylogenetic analyses were performed. All char-acterized enzymes involved in the biosynthesis ofisoprenoid side-chains and the glycerol phosphatebackbone along with their assembly in ether lipidswere included in these analyses. The sequence dataavailable in public databases was complemented byan in-depth sampling of isoprenoid lipid biosynthesisgenes from multiple genera of the archaeal orderHalobacteriales, allowing us to look at the evolutionof these enzymes on a smaller phylogenetic scale.This investigation of the isoprenoid biosynthesisapparatus of archaea on small and large phylogeneticscales reveals that it evolved through a combinationof evolutionary processes, including the co-optionof ancestral enzymes, modification of enzymaticspecificity, orthologous and non-orthologous genedisplacement, integration of components fromeukaryotes and bacteria and lateral gene transferwithin and between archaeal orders.
Introduction
Archaeal membrane lipids have several interesting char-
acteristics which distinguish them from their bacterial andeukaryotic counterparts: (i) isoprenoid, not fatty acids,side-chains; (ii) ether, not ester, links joining these side-chains to the glycerol phosphate backbone; and (iii) the
sn
-1, not
sn
-3, stereochemistry of this backbone.Archaeal lipids side-chains, like all other isoprenoids, areassembled from two universal precursors: isopentenyldiphosphate (IPP) and its isomer dimethylallyl diphos-phate (DMAPP). Most eukaryotes and some bacteria syn-thesize these precursors through the mevalonate pathway(Boucher and Doolittle, 2000; Lange
et al
., 2000). Thispathway has been described in details in these organismsand is always composed of five steps, as illustrated inFig. 1: (i) conversion of acetyl-CoA and acetoacetyl-CoAto 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA); (ii) reduc-tion of HMG-CoA to mevalonate; (iii) phosphorylation ofmevalonate; (iv) phosphorylation of phosphomevalonate;and (v) conversion of diphosphomevalonate to IPP.Archaea also derive their isoprenoids from mevalonate,as shown by tracer studies more than two decades ago(Kates and Kushwaha, 1978). Orthologues of theenzymes catalysing the first three steps of the pathway inbacteria and eukaryotes can also be found in archaea(Smit and Mushegian, 2000). However, the last twoenzymes of the standard bacterial/eukaryotic mevalonatepathway (phosphomevalonate kinase or PMK and diphos-phomevalonate decarboxylase or PPMD) are missing inmost archaea. Only one exception to this is known, whichis the identification of PPMD gene in the genome of theextremely halophilic archaeon
Halobacterium
sp. NRC-1(Ng
et al
., 2000). A type 1 isopentenyl diphosphateisomerase (IDI1), the enzyme responsible for the conver-sion of IPP to DMAPP in most eukaryotes and bacteriabut generally absent from archaea, was also found in thisgenome. In some archaea, this enzymatic reaction couldbe catalysed by a functional analogue of IDI1, the non-homologous type 2 isopentenyl diphosphate isomerase(IDI2). Kaneda
et al
. (2001) have shown IDI2 to be partof the mevalonate pathway gene cluster of the actino-mycete
Kitasatospora griseola
and identified its presencein several other bacteria and archaea.
The products of the mevalonate pathway (IPP andDMAPP) are used by archaea as building blocks for lipidside-chains. In acyclic lipids, these side-chains are com-posed of 20 or 25 carbons (C20 or C25). Such a lengthis reached through the sequential condensation of IPP

516
Y. Boucher, M. Kamekura and W. F. Doolittle
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
52
, 515–527
to a growing allylic polyisoprenoid diphosphate (Fig. 1).The first molecule in the chain is the IPP isomerDMAPP, to which IPP molecules are successively addedto obtain GPP (geranyl diphosphate, 10 carbons), FPP(farnesyl diphosphate, 15 carbons), GGPP (geranylgera-nyl diphosphate, 20 carbons) and FGPP (farnesylgera-nyl diphosphate, 25 carbons) (Fig. 1). These chainelongation reactions are catalysed by isoprenyl diphos-phate synthases, such as GGPP and FGPP synthases,which can differ from one another by the allylic sub-strate they accept to start the elongation process andthe chain length of their product(s) (Kellogg and Poulter,1997). In archaea, GGPP synthase can elongateDMAPP to obtain both FPP and GGPP, the latter beingthe isoprenyl forming the side-chains of C20–C20diether lipids. Archaea harbouring C20–C25 or C25–C25 diether lipids require an FGGP synthase, which caneither elongate directly from DMAPP [i.e.
Aeropyrumpernix
, see (Tachibana
et al
., 2000)] or from longer
allylic substrates like GGPP [
Natronobacterium
, see(Tachibana, 1994)].
The sequence of the enzyme responsible for linking thefirst side-chain to the glycerol phosphate backbone ofC20–C20 diether lipids was recently obtained from
Meth-anothermobacter thermoautotrophicus
(Soderberg
et al
.,2001). This enzyme, termed geranylgeranylglyceryl phos-phate (GGGP) synthase, was identified in six otherarchaeal species by similarity searches (Soderberg
et al
.,2001). It strongly favours
sn
-glycerol-1-phosphate as asubstrate, and therefore plays a role in defining the stere-oconfiguration of archaeal lipids. The enzyme responsiblefor adding the second C20 isoprenoid side-chain in
M.thermoautotrophicus
diether lipids, digeranylgeranylglyc-eryl phosphate (DGGGP) synthase, was identified in acellular fraction separate from GGGP synthase, but itsamino acid sequence has yet to be determined (Zhangand Poulter, 1993).
For a long time after the unique stereoconfiguration of
GGPP synthase
DMAPPIPP
GPP
FPP
GGPP
sn-Glycerol-1-P
DHAP
Glycerol-1-Pdehydrogenase
Glycerol-3-Pdehydrogenase
Eukaryotic / Bacterial ether and ester lipids
GGGPsynthase
DGGGPsynthase
sn-Glycerol-3-P
O P PO P P
O P P
O P P
O P P
O P P
FGPPsynthase
FGPP
OPO
O
OPOH
O
H
H
OOH
H
OPOH
H
P
OH OH
OPOH
+
GGGP
DGGGP (C20-C20 diether lipid)
C20-C25 diether lipid
C25-C25 diether lipid
Acetyl-CoA
HMG-CoA synthase
HMG-CoAreductase
HMG-CoA
Mevalonate
Mevalonatekinase (MK)
Mevalonate-5-P
Phospho-mevalonatekinase (PMK)
(HMGR)
(HMGS)
Mevalonate-5-PP
IPP isomerase
Mevalonate-5-PP decarboxylase (PPMD)
O P POH
OHO
O POH
OHO
OH
OHO
OH
OH
OHO
SCoA
O
SCoA
O O
SCoA
Acetoacetyl-CoA Central metabolism
A
A
A
A
S
S
S
S
O
S
FGPP synthaseor
GGPP synthase
FGPP synthaseor
GGPP synthase
FGPP synthaseor
S
IDI1
IDI2 AS
S
(G3PD) (G1PD)
Fig. 1.
Pathways for the synthesis of isoprenoid lipids and their building blocks. A boxed ‘A
¢
’ indicates that an enzyme is present in all archaea and a boxed ‘S
¢
’ indicates that it is present in only a subset of them. Abbreviations: phosphate (P), isopentenyl diphosphate (IPP), dimethylallyl diphosphate (DMAPP), GPP (ger-anyl diphosphate), FPP (farnesyl diphosphate), GGPP (geranylgeranyl diphosphate), FGPP (farnesylgeranyl diphosphate), GGGP (geranylgeranylglyceryl phosphate), DGGGP (digeranylgeranylglyceryl phosphate).

Isoprenoid lipid biosynthesis in archaea
517
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
52
, 515–527
archaeal lipids was determined, there have been ques-tions about the precursor used to synthesize the glycerolphosphate backbone. Nishihara and Koga (1995) identi-fied
sn
-glycerol-1-phosphate (G1P) dehydrogenase tobe responsible for the synthesis of the
sn
-glycerol-1-phosphate phospholipid backbone from dihydroxyacetonephosphate (DHAP) in
M. thermoautotrophicus
. They sub-sequently demonstrated that the activity specific to thisenzyme occurred in five other archaeal species and thatthe gene encoding for this enzyme was present in allcomplete archaeal genomes available at the time (Nishi-hara
et al
., 1999).These recent advances on the biochemistry of enzy-
matic steps leading to the biosynthesis of archaeal lipidsneed to be complemented by a better understanding oftheir origins and evolution. To this end, we performedphylogenetic analyses on all characterized enzymesinvolved in any steps of archaeal isoprenoid lipid biosyn-thesis, from the initial reaction of the mevalonate pathwayto the linkage between the glycerol phosphate backboneand the first isoprenoid side-chain. Because data avail-able from public databases was insufficient for evolution-ary analysis on a smaller phylogenetic scale, the genesencoding four isoprenoid biosynthesic enzymes (PPMD,IDI1, IDI2 and GGGP synthase) were amplified by PCRfrom several genera of the extremely halophilic archaealorder Halobacteriales.
Results and discussion
Biosynthesis of isoprenoid building blocks: IPP and DMAPP
Some components of the mevalonate pathway areconserved across the domains of life.
All organisms har-bouring a functional mevalonate pathway possess homol-ogous enzymes catalysing the first three steps: HMG-CoAsynthase (HMGS), HMG-CoA reductase (HMGR) andmevalonate kinase (MVK). As a rule, Life’s three domains(Bacteria, Archaea and Eukarya) exhibit monophyly forthese enzymes (all bacterial versions being more likeeach other, and so forth), but there are some striking andwell-supported exceptions. The HMGS gene of the greennon-sulphur bacterium
Chloroflexus aurantiacus
showshigh similarity to homologues from archaea, suggestingthat it was acquired from the latter group by lateral genetransfer (LGT) (Fig. 2A). A high divergence of haloar-chaeal HMGSs from other archaeal homologues is alsoobserved, but is most likely caused by the biased aminoacid composition typical of the proteins of extremely halo-philic archaea (Dennis and Shimmin, 1997).
Other deviations from the general monophyly of thearchaeal orthologues catalysing the first three steps of themevalonate pathway are the proteobacteria
Vibrio chol-
erae
and
Paracoccus zeaxanthificans
, as well as actino-mycetes species of the genera
Streptomyces
and
Kitasatospora
, which have acquired their HMGR fromarchaea [Fig. 2B, also see (Boucher and Doolittle, 2000)].Some archaea are also known to display mevalonatepathway genes acquired by LGT, as all representatives ofthe euryarchaeal orders Archaeoglobales and Thermo-plasmatales harbour an HMGR gene of seemingly bacte-rial origin (Boucher
et al
., 2001).In bacteria and fungi, the step of the mevalonate path-
way following the phosphorylation of mevalonate by MVKis catalysed by a homologue of this enzyme, phosphom-evalonate kinase (PMK). Eukaryotes belonging to theplant and animal kingdoms use a kinase showing littlesequence similarity to the bacterial/fungal PMKs to phos-phorylate phosphomevalonate. Previous to this study, nohomologues of either of these functionally analogousPMK enzymes had been found in archaea.
The full complement of mevalonate pathway enzymes are found in Sulfolobus
Our database survey revealed that both
Sulfolobus
species of which the genome has been completelysequenced (
S. tokadaii
and
S. solfataricus
) harbour aprotein with a high degree of similarity to the PMK foundin fungi and bacteria. Phylogenetic analysis of MVK andPMK (which are homologous enzymes) cluster this
Sul-folobus
protein with PMKs from the fungi
Schizosaccha-romyces pombe
and
Saccharomyces cerevisiae
(Fig. 2C).A protein showing high similarity to PPMD from eukary-
otes is encoded adjacent to this putative PMK in thegenomes of
S. tokadaii
and
S. solfataricus.
In phyloge-netic analysis, this protein clusters very strongly witheukaryotic homologues (Fig. 2D). This suggests that thisPPMD homologue was acquired from eukaryotes by anancestral
Sulfolobus
. The same is likely to hold for PMK,as this enzyme is not found in any other archaea and the
Sulfolobus
homologues show some affinity to PMKs fromfungi. The presence of these two enzymes in
Sulfolobus
means that this is the only genus of archaea known topossess all five enzymes of the standard mevalonatepathway.
Acquisition of mevalonate pathway enzymes from bacteria is an ancestral feature of extremely halophilic archaea
Sulfolobus
are not the only archaea to harbour a PPMD.The complete genome sequence of
Halobacterium
sp.NRC-1 also reveals the presence of a PPMD homologue(Ng
et al
., 2000). Our database survey detected thisenzyme in the three partially or completely sequencedThermoplasmatales genomes (
Thermoplasma acido-philum
,
Thermoplasma volcanium
and
Ferroplasma
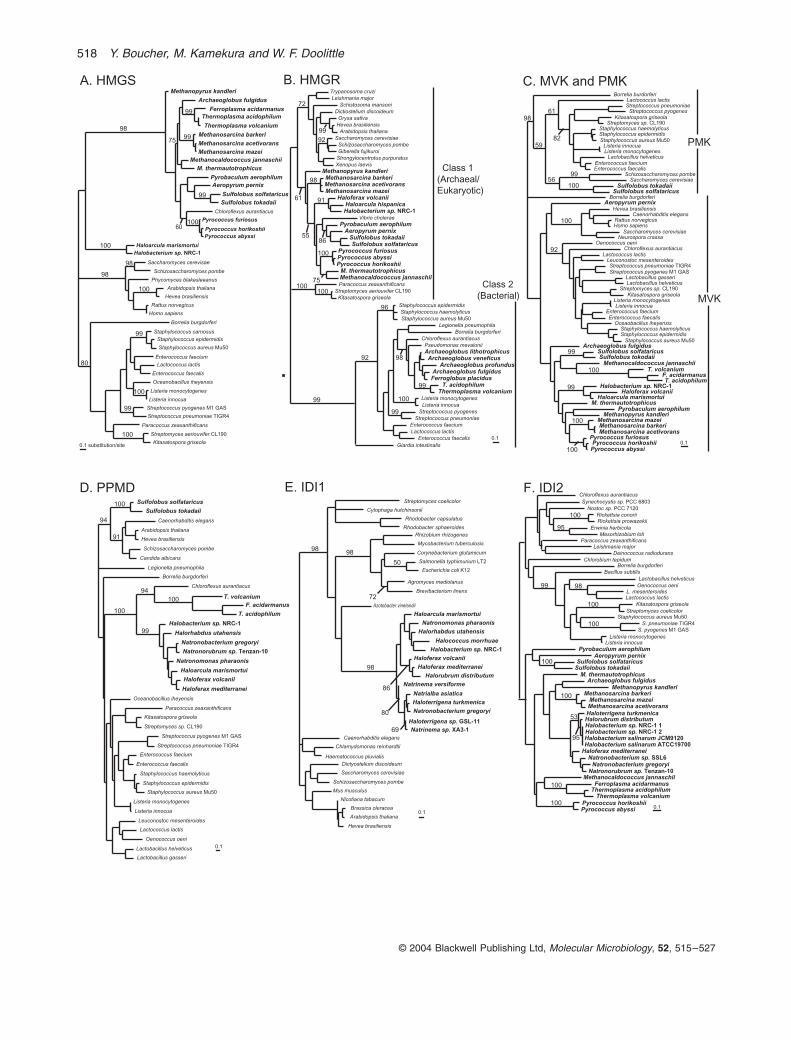
518
Y. Boucher, M. Kamekura and W. F. Doolittle
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
52
, 515–527
Trypanosoma cruziLeishmania major
Schistosoma mansoniDictiostelium discoideum
Orysa sativaHevea brasiliensis
Arabidopsis thalianaSaccharomyces cerevisiae
Schizosaccharomyces pombeGiberella fujikuroi
Strongylocentrotus purpuratusXenopus laevis
Methanopyrus kandleriMethanosarcina barkeriMethanosarcina acetivoransMethanosarcina mazei
Haloferax volcaniiHaloarcula hispanica Halobacterium sp. NRC-1
Vibrio choleraePyrobaculum aerophilum
Aeropyrum pernixSulfolobus tokadaiiSulfolobus solfataricus
Pyrococcus furiosusPyrococcus abyssiPyrococcus horikoshii
M. thermautotrophicusMethanocaldococcus jannaschii
Giardia intestinalis
Staphylococcus epidermidisStaphylococcus haemolyticusStaphylococcus aureus Mu50
Legionella pneumophilaBorrelia burgdorferi
Chloroflexus aurantiacusPseudomonas mevaloniiArchaeoglobus lithotrophicusArchaeoglobus veneficus
Archaeoglobus profundusArchaeoglobus fulgidusFerroglobus placidus
T. acidophilumThermoplasma volcanium
Listeria monocytogenesListeria innocua
Streptococcus pyogenes Streptococcus pneumoniae
Enterococcus faecium Lactococcus lactis
Enterococcus faecalis
Paracoccus zeaxanthificansStreptomyces aeriouvifer CL190
Kitasatospora griseola
99
92
96
100
100
99
99
98
100
98
100
75
86
91
55
61
72
9992
0.1
Class 2(Bacterial)
Class 1(Archaeal/Eukaryotic)
B. HMGR
Aeropyrum pernixPyrobaculum aerophilum
Methanopyrus kandleriArchaeoglobus fulgidus
Methanosarcina barkeri
Methanosarcina mazeiMethanosarcina acetivorans
M. thermautotrophicusMethanocaldococcus jannaschii
Ferroplasma acidarmanus
Thermoplasma volcanium
Thermoplasma acidophilum
Chloroflexus aurantiacus
Pyrococcus furiosus
Pyrococcus abyssiPyrococcus horikoshii
Sulfolobus solfataricusSulfolobus tokadaii
Borrelia burgdorferi
Haloarcula marismortuiHalobacterium sp. NRC-1
Phycomyces blakesleeanus
Saccharomyces cerevisae
Schizosaccharomyces pombe
Rattus norvegicus
Homo sapiens
Hevea brasiliensis
Arabidopsis thaliana
Streptococcus pyogenes M1 GAS
Streptococcus pneumoniae TIGR4
Oceanobacillus iheyensis
Listeria monocytogenes
Staphylococcus carnosusStaphylococcus epidermidis
Staphylococcus aureus Mu50
Enterococcus faeciumLactococcus lactis
Enterococcus faecalis
Paracoccus zeaxanthificans
Streptomyces aeriouvifer CL190
Kitasatospora griseola
Listeria innocua
98
99
99
10060
75
99
100
98
100
98
80
100
99
99
100
0.1 substitution/site
A. HMGSBorrelia burdorferi
Lactococcus lactisStreptococcus pneumoniae
Streptococcus pyogenesKitasatospora griseola
Streptomyces sp. CL190Staphylococcus haemolyticusStaphylococcus epidermidisStaphylococcus aureus Mu50
Listeria innocuaListeria monocytogenes
Lactobacillus helveticusEnterococcus faeciumEnterococcus faecalis
Schizosaccharomyces pombeSaccharomyces cerevisiae
Sulfolobus tokadaiiSulfolobus solfataricus
Borrelia burgdorferi Aeropyrum pernix
Hevea brasiliensis Caenorhabditis elegans
Rattus norvegicus Homo sapiens
Saccharomyces cerevisiaeNeurospora crassa
Oenococcus oeniChloroflexus aurantiacus
Lactococcus lactis Leuconostoc mesenteroidesStreptococcus pneumoniae TIGR4Streptococcus pyogenes M1 GAS
Lactobacillus gasseriLactobacillus helveticus
Streptomyces sp. CL190Kitasatospora griseola
Listeria monocytogenesListeria innocua
Enterococcus faeciumEnterococcus faecalis
Oceaobacillus iheyensisStaphylococcus haemolyticusStaphylococcus epidermidis
Staphylococcus aureus Mu50Archaeoglobus fulgidus
Sulfolobus solfataricus Sulfolobus tokodaii
Methanocaldococcus jannaschiiT. volcanium
F. acidarmanusT. acidophilum
Halobacterium sp. NRC-1Haloferax volcanii
Haloarcula marismortuiM. thermautotrophicus
Pyrobaculum aerophilum Methanopyrus kandleri
Methanosarcina mazeiMethanosarcina barkeriMethanosarcina acetivorans
Pyrococcus furiosusPyrococcus horikoshii
Pyrococcus abyssi
98
99
92
10056
59
100
61
82
100
100
99
99
100
0.1
C. MVK and PMK
MVK
PMK
Sulfolobus solfataricus
Sulfolobus tokadaii
Caenorhabditis elegans
Arabidopsis thaliana
Hevea brasiliensis
Schizosaccharomyces pombe
Candida albicans
Legionella pneumophila
Borrelia burgdorferi
Chloroflexus aurantiacus
T. volcaniumF. acidarmanus
T. acidophilum
Halobacterium sp. NRC-1
Halorhabdus utahensis
Natronobacterium gregoryi
Natronorubrum sp. Tenzan-10
Natronomonas pharaonis
Haloarcula marismortui
Haloferax volcanii
Haloferax mediterranei
Oceanobacillus iheyensis
Paracoccus zeaxanthificans
Kitasatospora griseola
Streptomyces sp. CL190
Streptococcus pyogenes M1 GAS
Streptococcus pneumoniae TIGR4
Enterococcus faecium
Enterococcus faecalis
Staphylococcus haemolyticus
Staphylococcus epidermidis
Staphylococcus aureus Mu50
Listeria monocytogenes
Listeria innocua
Leuconostoc mesenteroides
Lactococcus lactis
Oenococcus oeni
Lactobacillus helveticus
Lactobacillus gasseri
100
94
91
100
99
94
100
0.1
D. PPMD
Pyrobaculum aerophilum
Methanocaldococcus jannaschii
M. thermautotrophicus
Pyrococcus horikoshiiPyrococcus abyssi
Aeropyrum pernix
Borrelia burgdorferi
Sulfolobus solfataricusSulfolobus tokadaii
Methanopyrus kandleriArchaeoglobus fulgidus
Methanosarcina barkeriMethanosarcina mazeiMethanosarcina acetivorans
Ferroplasma acidarmanusThermoplasma acidophilum
Thermoplasma volcanium
Haloferax mediterranei
Haloterrigena turkmenicaHalorubrum distributumHalobacterium sp. NRC-1 1Halobacterium sp. NRC-1 2Halobacterium salinarum JCM9120Halobacterium salinarum ATCC19700
Natronobacterium gregoryiNatronobacterium sp. SSL6
Nostoc sp. PCC 7120
Chloroflexus aurantiacusSynechocystis sp. PCC 6803
Rickettsia conoriiRickettsia prowazekii
Mesorhizobium lotiErwinia herbicola
Chlorobium tepidum
Paracoccus zeaxanthificans
Deinococcus radioduransLeishmania major
Kitasatospora griseola Streptomyces coelicolor
Staphylococcus aureus Mu50
S. pyogenes M1 GASS. pneumoniae TIGR4
Lactobacillus helveticusOenococcus oeni
Lactococcus lactisL. mesenteroides
Listeria monocytogenesListeria innocua
Bacillus subtilis
100
100
100
100
100
100
100
99 98
95
F. IDI2
Natronorubrum sp. Tenzan-10
53
95
0.1
Streptomyces coelicolor
Cytophaga hutchinsonii
Rhodobacter capsulatus
Rhodobacter sphaeroidesRhizobium rhizogenes
Mycobacterium tuberculosis
Corynebacterium glutamicum
Salmonella typhimurium LT2
Escherichia coli K12
Agromyces mediolanus
Brevibacterium linens
Azotobacter vinelandii
Haloarcula marismortuiNatronomonas pharaonis
Halorhabdus utahensis
Halococcus morrhuaeHalobacterium sp. NRC-1
Haloferax volcaniiHaloferax mediterranei
Halorubrum distributumNatrinema versiforme
Natrialba asiaticaHaloterrigena turkmenicaNatronobacterium gregoryi
Haloterrigena sp. GSL-11Natrinema sp. XA3-1
Caenorhabditis elegans
Chlamydomonas reinhardtii
Haematococcus pluvialisDictyostelium discoideum
Saccharomyces cerevisiae
Schizosaccharomyces pombe
Mus musculus
Nicotiana tabacum
Brassica oleracea
Arabidopsis thaliana
Hevea brasiliensis
98 98
98
72
50
86
80
69
0.1
E. IDI1

Isoprenoid lipid biosynthesis in archaea
519
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
52
, 515–527
acidarmanus
). However, phylogenetic analysis indicatesthat the PPMD found in
Halobacterium
and the Thermo-plasmatales are likely to have a different origin than the
Sulfolobus
enzyme. Indeed, these PPMD genes clusterwith bacterial homologues and are distinct from eukaryoticsequences (Fig. 2D). Like its HMGS gene,
Chloroflexusaurantiacus
PPMD clusters strongly with archaealsequences and could have originated from Thermoplas-matales or extremely halophilic archaea (Fig. 2D). Alter-natively,
Chloroflexus
could be the source of the PPMDfound in these archaea. This second alternative seemsless likely, as
Chloroflexus
PPMD gene clusters strongly
in-between the Halobacteriales and Thermoplasmataleshomologues in phylogenetic analyses, as opposed tooccupying a basal position if it was the ancestralsequence.
Another isoprenoid biosynthesis enzyme usuallymissing from archaea, type 1 isopentenyl diphosphateisomerase (IDI1), was found in the genome of
Halobacte-rium
sp. NRC-1 (Ng
et al
., 2000). This enzyme was pre-viously known only from eukaryotes and bacteria. Ourdatabase survey revealed that all archaea, including
Halobacterium
, harbour the analogous type 2 IDI (IDI2)described by Kaneda
et al
. (2001). This alternative IPP
Fig. 2.
Best maximum likelihood phylogenetic trees for various enzymes involved in the biosynthesis of isoprenoid lipids in archaea. Bootstrap support values correspond to the consensus of 100 Fitch–Margoliash maximum likelihood distance trees. Trees are arbitrarily rooted and only relevant nodes with support values over 50% are displayed. Archaeal taxa are highlighted in bold.A. HMG-CoA synthase (HMGS).B. HMG-CoA reductase (HMGR) rooted with class 2 enzymes.C. Phosphomevalonate kinase (PMK) rooted with Mevalonate kinase (MVK).D. Diphosphomevalonate decarboxylase (PPMD).E. Isopentenyl diphosphate isomerase type 1 (IDI1).F. Isopentenyl diphosphate isomerase type 2 (IDI2).G. Short (10–25 carbons) and medium (30–50 carbons) chain isoprenyl diphosphate synthases. All archaeal short-chain enzymes are geranylger-anyl diphosphate (GGPP, 20 carbons) synthases, except for the
Aeropyrum pernix
homologue, which is a farnesylgeranyl diphosphate (FGPP, 25 carbons) synthase. The residues found at positions 74 and 77 in the amino acid sequence of each taxa (positions based on
Sulfolobus acidocaldarius
GGPP synthase) are indicated in parenthesis beside the taxa names. These residues are believed to be important in limiting the length of the isoprenyl diphosphate produced (see
Text
).H.
sn
-Glycerol-1-phosphate dehydrogenase (G1PD) rooted with glycerol dehydrogenase (GD).I. Geranylgeranylglyceryl phosphate (GGGP) synthase.
Streptomyces coelicolorMycobacterium tuberculosis
Pasteurella multocidaRalstonia solanacearum
Pseudomonas aeruginosaNostoc sp. PCC 7120
Capsicum annuumArabidopsis thalianaMentha piperita
Chlorobium tepidum Caulobacter crescentusBrucella melitensisRhodospirillum rubrum
Paracoccus zeaxanthificansRhodobacter sphaeroides
Xanthomonas campestrisMicrobulbifer degradansPseudomonas aeruginosa
Pseudomonas sp. BG33RPasteurella multocida
Escherichia coli K12Yersinia pestisNeisseria meningitidisNitrosomonas europea
Ralstonia solanacearumOceanobacillus iheyensis
Bacillus stearothermophilusCyanidium caldarium
Porphyra purpureaNostoc sp. PCC 7120
Prochlorococcus marinusSynechocystis sp. PCC 6803
Arabidopsis thalianaSchizosaccharomyces pombe
Saccharomyces cerevisiaeHomo sapiens
Drosophila melonagasterAquifex aeolicus
Thermoplasma volcaniumThermoplasma acidophilum
P. aerophilum Sulfolobus tokodaii
S. solfataricusArchaeoglobus fulgidusMethanosarcina acetivoransMethanosarcina mazei
Archaeoglobus fulgidus
Halobacterium sp. NRC-1Methanosarcina acetivorans
Methanosarcina mazei
Methanopyrus kandleriMethanothermobacter marburgensis
M. thermautotrophicusMethanocaldococcus jannaschii
Pyrobaculum aerophilumPyrococcus horikoshiiPyrococcus abyssi
Sulfolobus solfataricusSulfolobus tokodaii
Thermoplasma volcaniumT. acidophilum
Aeropyrum pernix
92
97 100
94
97
99
98
100
98(+indel)
100
80
10059
50
57
50
100
100
100
100
10083
100
100
(LA)
(LA)
(LS)
(IS)
(VA)
(IA)(IA)
(IA)(IA)
(IA)(IA)
(IA)
(IA)
(IA)(IA)
(IA)
(IA)(IA)
(IA)(IA)
(IA)(IA)
(IA)(IA)
(IA)(IA)
(IA)(IA)
(IA)(IA)
(IA)
(LA)(LA)
(LV)
(IA)(MA)
(MA)(VF)
(VF)(VY)
(IY)(IY)
(IM)(IM)
(IM)(IM)
(IF)(VF)
(VF)(IF)
(IF)(IF)
(IF)(IY)
(IY)(IY)
(IY)(LF)
(LF)(AY)
(AY)(AF)
Medium-chainisoprenyl diphosphatesynthases
Short-chainisoprenyl diphosphatesynthases(Bacterial and PlantsGGPP synthases)
Short-chainisoprenyl diphosphatesynthases(Archaeal FGPP/GGPP synthases)
0.1 substitution/site
G. Isoprenyl diphosphate I. GGGP synthase
0.1
synthases
Cytophaga hutchinsonii
Natronobacterium gregoryi 1
Haloterrigena turkmenica
Haloferax volcanii 1
Halorubrum distributum
Halobacterium sp. NRC-1 1
Halococcus morrhuae
Haloferax volcanii 2
Halobacterium sp.NRC-1 2
Natronobacterium
Natrialba asiatica
Archaeoglobus fulgidus
Staphylococcus aureus Mu50
Oceanobacillus iheyensis
Bacillus halodurans
Bacillus subtilis
Bacillus anthracis
Listeria innocua
Listeria monocytogenes
Sulfolobus solfataricus
Sulfolobus tokodaii
Aeropyrum pernix
Pyrobaculum aerophilum
Methanopyrus kandleri
M. thermautotrophicus
Methanocaldococcus jannaschii
Thermoplasma volcanium
Thermoplasma acidophilum
Methanosarcina acetivorans
Methanosarcina mazei
Methanosarcina barkeri
Pyrococcus horikoshii
Pyrococcus abyssi
Pyrococcus furiosus
gregoryi 2
Haloarcula marismortui100(+2 indels)
98
90
86
100
63
86
100
80
91
79
100
100
98
Nostoc sp. PCC 7120
Sinorhizobium meliloti
Thermotoga maritima
Salmonella typhimurium LT2 1
Clostridium perfringens
Oceanobacillus iheyensis
Geobacillus stearothermophilus
Schizosaccharomyces pombe
Streptococcus pneumoniae TIGR4
Streptococcus pyogenes M1 GAS
Thermobifida fusca
Halobacterium sp. NRC-1
Halococcus morrhuae
Natrialba asiatica
Citrobacter freundii
Salmonella typhimurium LT2 2
Escherichia coli K12
Pyrobaculum aerophilum
Aeropyrum pernix
Sulfolobus tokodaii
Sulfolobus solfataricus
Archaeoglobus fulgidus
Methanopyrus kandleri
Pyrococcus furiosus
Pyrococcus abyssi
Pyrococcus horikoshii
M. thermautotrophicus
Methanosarcina barkeri
Methanosarcina mazei
Methanosarcina acetivorans
Haloferax volcanii
Halobacterium sp. NRC-1
Haloarcula marismortui
Ferroplasma acidarmanus
Thermoplasma acidophilum
Thermoplasma volcanium
Methanococcus jannaschii
98
88
94
88 98
96
100
GD
G1PD
0.1
H. GD and G1PD

520
Y. Boucher, M. Kamekura and W. F. Doolittle
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
52
, 515–527
isomerase, first discovered by these workers in the meva-lonate gene cluster of
Kitasatospora griseola
, does notshare detectable sequence similarity with the bacterial/eukaryotic IDI1. In addition to archaea and
Kita-satospora
, IDI2 is found in various bacteria (althoughonly those that do not possess IDI1) and in the trypano-somatid
Leishmania major
. An IDI is essential to allorganisms possessing only the mevalonate pathway tosynthesize their isoprenoids, as they need the enzyme tomake the essential precursor DMAPP in addition to IPP.This requirement is satisfied by the presence of an IDI ofeither type in eukaryotes and bacteria, and of an IDI2 inall archaea except extremely halophilic archaea, whichare so far the only organisms known to harbour bothenzymes.
To confirm that the presence of a PPMD and two typesof IDIs was a general trait of the Halobacteriales (thearchaeal order representing extreme halophiles) and notsimply a specific feature of
Halobacterium
, the genesencoding these three enzymes were PCR amplified fromrepresentatives of several genera of this order. Table 1reports the results of this survey, in which PPMD wasobtained from four species, IDI1 from seven and IDI2 fromthree. PPMD and IDI1 have also been identified in the twohaloarchaea for which a genome sequencing project isunder way (
Haloferax volcanii
and
Haloarcula marismor-tui
), whereas IDI2 has been detected in one of the two(
Haloferax volcanii
) (Table 1).The fact that these genes are short and present rela-
tively few highly conserved regions makes the design ofefficient PCR primers difficult. Failure to consistentlyamplify these genes from the DNA of all strains on whichPCR was performed should therefore not be interpretedas the absence of the gene in question. The high num-bers of strains from which each of these genes wasnonetheless amplified does suggest that they are a uni-versal feature of Halobacteriales, or at least present inthe vast majority of species in this group. In phylogeneticanalyses of all three genes (Fig. 2D–F), Halobacterialesform monophyletic clusters, suggesting that they areancestrally present in this order. The distribution and phy-logeny of IDI1 and PPMD suggest that they wereacquired by LGT from bacteria. Both of these enzymesare found in several groups of bacteria and eukaryotes,but are otherwise very limited in their distribution inarchaea. IDI1 is found only in Halobacteriales, whereasPPMD is also found in Thermoplasmatales and
Sulfolo-bus
, although the homologues found in the latter grouphave a different (eukaryotic) origin. Also, each of thesetwo enzymes clusters with bacteria in phylogenies, albeitweakly.
For its part, IDI2 does not seem to have been involvedin interdomain LGT. It is present in all archaea, whichform a monophyletic cluster that includes the Halobacte-
riales orthologues. Despite the apparent absence ofinterdomain lateral transfer, there seems to have beenexchange of this gene between species of Halobacteri-ales. Phylogenies of the different isoprenoid lipid biosyn-thesis genes amplified from a variety of Halobacterialesare mostly unresolved, as these genes are short and onlypartial sequences were obtained for most of them. How-ever, there are two clades of Haloarchaea found in mostof these phylogenies as well as in a phylogeny of thesmall ribosomal subunit (SSU) rRNA gene: one includesmostly neutrophilic halophiles (subgroup I) and the othercontains mostly haloalkaliphiles (subgroup II) (Fig. 3).This division does not hold for IDI2, as
Haloterrigenaturkmenica
, clustering strongly with the subgroup II in theSSU and IDI1 phylogenies, clusters just as strongly withsubgroup I in the IDI2 phylogeny (Fig. 3). This suggestsLGT between genera of Halobacteriales for the IDI2gene.
Biosynthesis of isoprenoid side-chains: isoprenyl diphosphate synthases
Functional plasticity of isoprenyl diphosphate synthases.
Mutational studies on the GGPP synthase of
Sulfolobusacidocaldarius
demonstrated the extent of the plasticity ofisoprenyl diphosphate synthases regarding the length ofthe isoprenoid products. A single amino acid substitutionchanged this archaeon GGPP synthase into an enzymesynthesizing FGPP as its main product and able to pro-duce small amounts of hexaprenyl diphosphate (30 car-bons isoprenyl diphosphate) (Ohnuma
et al
., 1997). Withtwo or three amino acid substitutions, using DMAPP, FPPor GPP as the allylic substrate, the main product couldreach lengths of 35 or 40 carbons (heptaprenyl and octa-prenyl diphosphates) and secondary products reachinglengths of up to 65–120 carbons were also obtained(Ohnuma
et al
., 1998).A naturally occurring example of such chain elongation
plasticity is found in
Aeropyrum pernix
. This archaeononly harbour C25–C25 diether lipids, a rare featureamong archaea, most of which produce only C20–C20lipids (De Rosa and Gambacorta, 1988). This hyperther-mophile does not possess a GGPP synthase, but a sin-gle FGPP synthase (Tachibana
et al
., 2000). The latterwas most likely derived from an ancestral archaealGGPP synthase, as suggested by a phylogenetic analy-sis performed by Tachibana
et al
. (2000). Our phylogenyof isoprenyl diphosphate synthases also support thisclaim. In the best maximum likelihood tree,
A. pernix
FGPP synthase is found grouping among archaealGGPP synthases (Fig. 2G). Beside
A. pernix
FGPP syn-thase,
S. acidocaldarius
GGPP synthase and
M. ther-moautotrophicus
GGPP synthase, which have beenbiochemically characterized in detail (Chen and Poulter,

Isoprenoid lipid biosynthesis in archaea
521
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
,
52
, 515–527
1993; Ohnuma
et al
., 1996; Tachibana
et al
., 2000),assessement of the chain length specificity of isoprenyldiphosphate synthases is based on the amino acid resi-due found at position 74 (amino acid positions of
S. aci-
docaldarius
GGPP synthase). GGPP synthases harbouran isoleucine or leucine residue at this position, asopposed to the alanine found in the
A. pernix
enzyme(Fig. 2G). Bulkier residues at this position have been
Table 1.
Isoprenoid lipid biosynthesis enzymes found in Archaea.

522
Y. Boucher, M. Kamekura and W. F. Doolittle
© 2004 Blackwell Publishing Ltd,
Molecular Microbiology
, 52, 515–527
shown to ‘floor’ the hydrophobic pocket that contains theisoprenoid chain during the elongation process, effec-tively limiting the length of the isoprenoid products(Ohnuma et al., 1998). Exceptions to this are the Ther-moplasma, which, like A. pernix, harbour an alanine atposition 74. However, the T. acidophilum and T. volca-nium enzymes are almost certainly GGPP synthases, asthese archaea are known to only contain C20–C20diether lipids (Shimada et al., 2002). This suggests thatother residues beside the one found at position 74 couldlimit the length of the isoprenoid chain produced.
Diversity of isoprenyl diphosphate synthases in archaea
Similarity searches for archaeal isoprenyl diphosphatesynthases also recovered enzymes predicted to synthe-size medium-chain isoprenoids (30–50 carbons). A char-acteristic distinguishing short and medium-chain isoprenyldiphosphate synthases is the residue found in position 77.Short-chain specific enzymes almost always display a
bulky residue at this position (phenylalanine or tyrosine)whereas enzymes producing medium-chain products usu-ally harbour a smaller residue (alanine, serine or valine).This residue, like position 74, is thought to be orientedtoward the channel in which the isoprenoid chain is elon-gated, limiting the length of the product (Ohnuma et al.,1998). Several archaeal homologues divergent from theclade containing characterized short-chain isoprenyldiphosphate synthases all display an alanine, serine orvaline residue at position 77 (Fig. 2G) and are thereforemost likely medium-chain isoprenyl phosphate synthases,possibly involved in the biosynthesis of respiratory quino-nes (Kellogg and Poulter, 1997). These medium-chainenzymes were detected in only some of the archaea thathave their genome completely sequenced (Fig. 2G). Con-sequently, the archaea in which this medium-chain isopre-nyl diphosphate synthase cannot be detected must use adistantly related homologous prenyltransferase or a func-tionally analogous enzyme to synthesize their essentialisoprenoids.
Halorhabdus utahensis
Natrinema sp. XA3-1
Natrialba asiatica
Haloterrigena turkmenica
0.1 substitution/site
0.1 Natronorubrum sp. Tenzan-10 0.1
Halorhabdus utahensis
Natrinema versiforme
Natrinema sp. XA3-1
0.1
Haloferax mediterranei
0.1
Haloferax volcanii 2
Halobacterium sp. NRC-1 2
Natrialba asiatica
Halorubrum distributum
Haloterrigena turkmenica
Halococcus morrhuae
Haloferax volcanii 1
Natronobacterium gregoryi 1
Halobacterium sp. NRC-1 1
Natronobacterium gregoryi 2
Halorubrum distributum
Haloferax mediterranei
Haloferax volcanii
Halococcus morrhuaeHalobacterium sp. NRC-1
Haloterrigena turkmenica
Natronobacterium gregoryi
Natrialba asiatica
Natronomonas pharaonis
Halobacterium sp. NRC-1
Halorhabdus utahensis
Natronobacterium gregoryi
Haloferax mediterranei
Haloferax volcanii
Natronomonas pharaonis
Haloferax mediterranei
Haloferax volcanii
Halorubrum distributum
Natronomonas pharaonis
Halococcus morrhuae
Natrinema versiforme
Natronorubrum sp. Tenzan-10
Natronobacterium gregoryi
Halobacterium sp. NRC-1 SSU
PPMD IDI1
IDI2 GGGPS
97/100
55/--
71/--
66/96
44/--
75/59
95/95
77/98
99/100
84/76
92/98
44/--55/92
39/--53/62
51/6553/--
72/--
60
10057
53
936867
96
Halorubrum distributum
Haloterrigena turkmenica
Halobacterium sp. NRC-1
Halobacterium salinarum JCM9120
Halobacterium salinarum ATCC19700
Natronobacterium gregoryi
Natronobacterium sp. SSL6
Natronorubrum sp. Tenzan-10
71/--
90/82
99/100
98/100
53/54
Halobacterialessubgroup I(dominated byneutrophiles)
Halobacterialessubgroup II(dominated byalkaliphiles)
GTR+I+G
K81uf+G HKY+I+G
TrN+G TVM+I+G
Fig. 3. Best maximum likelihood phylogenetic tree for various enzymes involved in the biosyn-thesis of isoprenoid lipids in extremely halo-philic archaea. The trees shown are from an analysis at the amino acid level. The bootstrap support values displayed before the dash corresponds to the consensus of 100 Fitch-Margoliash maximum likelihood distance trees (amino acid analysis) and the value displayed after the dash corresponds to the consensus of 100 maximum likelihood trees (DNA analysis). If (–) is displayed instead of a bootstrap value, it means that the bootstrap consensus tree dis-agreed with the best tree or was not resolved at that node. The top tree was constructed from an alignment of small ribosomal subunit rRNA genes (SSU). Because this gene is not protein coding, only the DNA analysis bootstrap value is presented. The nucleotide substitution model used for the DNA analysis of each gene is indicated under its respective tree (I = invariant sites, G = gamma distributed rates).

Isoprenoid lipid biosynthesis in archaea 523
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 515–527
Biosynthesis of the glycerol phosphate backbone and its association with side-chains: sn-glycerol-1-phosphate dehydrogenase and geranylgeranylglyceryl phosphate synthase
Evolution of the stereoconfiguration of archaeal lipids.Two enzymes involved in the biosynthesis of archaeallipids show stereospecificity. sn-Glycerol-1-phosphate(G1P) dehydrogenase introduces stereospecificity intoarchaeal lipids by specifically synthesizing glycerol phos-phate with the sn-1 stereoconfiguration from dihydroxyac-etone phosphate (DHAP) (Nishihara and Koga, 1995).Geranylgeranylglyceryl phosphate (GGGP) synthasestrongly favours this glycerol phosphate stereoisomerwhen attaching the first isoprenoid side-chain, yielding 3-O-geranylgeranyl-sn-glycerol-1-phosphate. The attach-ment of the second side-chain is also stereospecific, asDGGGP synthase will only recognize the sn-1 phosphatemonoether as opposed to an sn-3 phosphate monoethersubstrate, yielding only archaeol (2,3-O-geranylgeranyl-sn-glycerol-1-phosphate) as a product (Zhang andPoulter, 1993). CDP-archaeol (the cytidylated version ofarchaeol) is thought to be a major phospholipid precursor(Morii et al., 2000). The enzyme responsible for its syn-thesis from CTP and archaeol, CDP-archaeol synthase,does not recognize the stereochemical structure of theglycerol phosphate backbone or the linkage betweenglycerol and the isoprenoid side-chains (ester or etherlinkage). It therefore seems that the specific stereo-configuration of archaeal lipids is established by G1Pdehydrogenase as well as the GGGP and DGGGPsynthases.
Origin of sn-glycerol-1-phosphate dehydrogenase
sn-Glycerol-1-phosphate dehydrogenase and glyceroldehydrogenase catalyse similar reactions, the substrateand product being phosphorylated in one case and not inthe other (dihydroxyacetone versus dihydroxyacetonephosphate as substrate and glycerol versus sn-glycerol-1-phosphate as product). These enzymes are alsohomologous, sharing about 20–25% identity in theiramino acid sequences. They are both part of the NAD-dependent dehydrogenase superfamily, which alsoincludes G3P dehydrogenases. Although the latterenzyme is functionally equivalent to G1P dehydrogenase(with the exception of its stereospecificity), they sharelittle sequence similarity.
Our database survey confirms earlier claims, based ona more limited sampling, that G1P dehydrogenase issolely found in archaea (Nishihara et al., 1999). Phyloge-netic analysis also presents G1P dehydrogenases as amonophyletic cluster among the larger NAD-dependentdehydrogenase superfamily (Fig. 2H). This suggests that
G1P dehydrogenase is an archaeal invention derived froman enzyme of the NAD-dependent dehydrogenase super-family, possibly glycerol dehydrogenase, which sharessimilar sequence, substrate and product. The latterenzyme is found only in the bacterial domain, with theexception of one eukaryote (Schizosaccharomycespombe) and haloarchaea, which both seem to haveacquired the enzyme by LGT (Fig. 2H).
sn-Glycerol-1-phosphate dehydrogenase could haveplayed a crucial role in the creation of a novel glycerolphosphate backbone stereospecificity and therefore thedifferentiation of archaea from their ancestors. However,the particular origin of this enzyme, which is likely to havebeen derived from the bacterial enzyme glycerol dehydro-genase, does not inform us on the origin of archaea them-selves. Indeed, before its co-option to a G1PD, glyceroldehydrogenase could have been obtained by proto-archaea either through vertical inheritance from theirdirect ancestor or by LGT from an already differentiatedbacterial domain.
Presence of two homologous types of GGGP synthases in archaea
The recently described sequence of the gene encodingfor Methanothermobacter thermoautotrophicus GGGPsynthase was the first obtained for this protein (Soderberget al., 2001). This enzyme is selective not only for the sn-glycerol-1-phosphate acceptor, but also for the isoprenoidside-chain added, strongly favouring GGPP over shorteror longer chains (Zhang and Poulter, 1993).
Our similarity searches reveal the presence of homo-logues of this enzyme in all archaea, even A. pernix,which, as mentioned earlier, is known to produce onlyC25–C25 diether lipids. Although M. thermoautotrophicusGGGP synthase shows very little activity with FGPP as aprenyl donor, the A. pernix homologue might be able touse this longer substrate, as this organism would havelittle use for a GGPP specific enzyme (Soderberg et al.,2001). Phylogenetic analysis shows the A. pernix homo-logue clustering among other archaeal enzymes whichare likely to display GGGP synthase activity, as the majorfraction of the lipids of these archaea are C20–C20diether lipids (Fig. 2I). This outlines that GGGP synthasehomologues might harbour a functional plasticity similarto other prenyltransferases; a few mutations could be suf-ficient to alter the length of the prenyl donor used by theenzyme.
A surprising finding from phylogenetic analysis is theexistence of two distinct but homologous types of GGGPsynthase. Halobacterium sp. NRC-1 and Archaeoglobusfulgidus both possess very divergent enzymes that clusterwith homologues from various species of the bacterialorder Bacillales. This separate cluster is not only sup-

524 Y. Boucher, M. Kamekura and W. F. Doolittle
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 515–527
ported by a high bootstrap value (100%, see Fig. 2I), butalso by two separate amino acid insertions found only inthe GGGP synthase orthologues that are part of this clus-ter (data not shown). To investigate if this divergentenzyme was a specific feature of Halobacterium or a moregeneral characteristic of the Halobacteriales, various gen-era of extremely halophilic archaea were surveyed for itspresence. In addition to the detection of its presence inthe three partially or completely sequenced haloarchaealgenomes, we obtained this divergent GGGP synthasefrom five genera of Halobacteriales (Table 1). In a fewspecies of haloarchaea, including Halobacterium, two par-alogues of this divergent enzyme were present (Fig. 2I).The role of these two paralogues is unclear, but it ispossible that one of them encodes a farnesylgeranylglyc-eryl phosphate synthase, as some Halobacteriales(among others Haloterrigena, Halococcus, Natronobacte-rium, Natrinema, Natrialba and Natronomonas) produceC20–C25 diether lipids (Kamekura and Kates, 1999).However, some Halobacteriales that only have C20–C20lipids (Haloferax and Halobacterium) also exhibit twoparalogues.
Given its presence in several Halobacteriales genera, adivergent type of GGGP synthase is most likely an ances-tral characteristic of this order. The situation could besimilar for the Archaeoglobales, but a survey of otherspecies in addition to Archaeoglobus fulgidus would berequired to confirm this. The evolutionary origin of thisdivergent type of GGGP synthase is unclear. The distri-bution of GGGP synthase homologues, ubiquitous inarchaea but only found in the order Bacillales and aCytophaga among bacteria, suggest that this enzyme isan archaeal invention, which was later acquired by bacte-ria through LGT. The Halobacteriales, Archaeoglobus andBacillales homologues are closely related to each otherand distinct from other archaeal homologues and theCytophaga gene. The Bacillales GGGP synthase homo-logues would therefore have originated from the Archaeo-globales or the Halobacteriales, whereas the Cytophagahomologue would descend from the enzyme of someother archaeal group. Since no ether lipids of the sn-2,3stereoconfiguration are found in bacteria, the enzymespresent in the Bacillales and Cytophaga were probablyco-opted for a different function. All Bacillales for whichsequence information is available harbour the GGGP syn-thase homologue, which is always encoded upstream ofa DNA helicase and an NAD-dependent DNA ligase. Thehomologue found in Staphylococcus aureus S20 (termedpcrB) has been identified as part of a chromosomaloperon that include the pcrA gene, which encodes a heli-case required for cell viability and the replication of plas-mid pT180 (Iordanescu, 1993). The ubiquity of this genein Bacillales and its linkage with pcrA points to an impor-tant function, possibly in DNA replication.
Lateral transfer of GGGP synthase homologues is alsolikely to have happened among archaea, as the specificrelationship found between Archaeoglobus and Halobac-teriales GGGP synthases (Fig. 2I) is never recoveredusing known phylogenetic markers (translational appara-tus proteins or the small ribosomal subunit gene) (Matte-Tailliez et al., 2002). What is found instead with mostmarkers is a clear monophyly of Halobacteriales and themethanogenic orders Methanosarcinales and Methanomi-crobiales (Matte-Tailliez et al., 2002). This evolutionaryrelationship between extreme halophiles and methano-gens is obviously not found in the GGGP synthase phy-logeny, where enzymes from the Methanosarcinales areclearly part of the main archaeal cluster (Fig. 2I). Thisincongruence between well-known phylogenetic markersand GGGP synthase suggests that the latter enzyme waslaterally transferred between the ancestors of the Halo-bacteriales and Archaeoglobus.
Concluding remarks
Biosynthesis of the isoprenoid building blocks IPP andDMAPP seems to be carried out through the same cata-lytic steps in all archaea, but with variability of theenzymes performing those reactions. The first threegenes of the mevalonate pathway are found in all archaea,with some lineages experiencing orthologous displace-ment from bacteria for the HMGR gene (Archaeoglobales,Thermoplasmatales). The homologues catalysing the lasttwo steps in bacteria and eukaryotes are only present ina few lineages of archaea. The lineages displaying thesegenes seem to have acquired them by LGT: from eukary-otes for the PMK and PPMD found in Sulfolobus and frombacteria for the PPMD found in Thermoplasmatales andHalobacteriales. Functional analogues of the bacterial/eukaryotic PMK and/or PPMD must be present in allarchaea in which one or both of these two enzymes can-not be found, as their function is absolutely essential. Thesubsequent conversion of the mevalonate pathway prod-uct IPP to its isomer DMAPP can also be catalysed bytwo analogous types of enzymes: one is found in allarchaea (type 2 IPP isomerase, IDI2) and the other is onlyfound in Halobacteriales (type 1 IPP isomerase, IDI1).This IDI1, found in all Halobacteriales examined, seemsto have been acquired by LGT from bacteria. Like all theenzymes of the mevalonate pathway of bacterial/eukary-otic origin, IDI1 is found in all representatives of thearchaeal order by which it was acquired. This makes itlikely that all LGT events so far identified to be responsibleof the presence of isoprenoid lipid biosynthesis enzymesin particular orders of archaea have occurred prior to thediversification of these groups.
These ancestral transfer events are not the only thingthat affected the evolution of isoprenoid lipid biosynthesis

Isoprenoid lipid biosynthesis in archaea 525
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 515–527
in archaea. Further LGT events within (IDI2) and between(GGGP synthase) archaeal orders must have influencedthe lipid metabolism of certain archaea. Also, the prenyl-transferases involved in the elongation of the isoprenoidside-chain (isoprenyl diphosphate synthases) and itsassociation to the glycerol phosphate backbone (GGGPsynthase) both seem to have experienced specificityswitches during archaeal evolution, an ancestral archaealGGPP synthase giving rise to the FGPP synthase foundin Aeropyrum pernix and an ancestral archaeal GGGPsynthase possibly giving rise to an FGGP synthase in thesame archaeon.
Most of the isoprenoid lipid biosynthesis enzymes seemto have been present before the divergence of the domainarchaea, as they are relatively widespread in otherdomains. Two of them, however, are likely to be archaealinventions: GGGP synthase and G1P dehydrogenase.Besides the GGGP synthase homologues found in thebacterial order Bacillales and a Cytophaga (likely to havebeen co-opted for a different function), these two enzymesare solely found in archaea and catalyse stereospecificreactions that have never been observed in the bacterialdomain. The type of enzyme the GGGP synthases origi-nated from is difficult to identify, as they do not sharesignificant similarity with any other prenyltransferases.However, G1P dehydrogenases are clearly part of theNAD-dependent dehydrogenase superfamily, and possi-bly originate from the co-option of glycerol dehydroge-nase, with which they share similar sequence, substrateand product.
Archaea display important variability in the polymerscomposing their cell envelope, with many lineage-specificinventions, such as pseudomurein in Methanobacteriales/Methanopyrales, methanochondroitin in Methanosarcinaor glutaminylglycan in Natronococcus (Kandler and Konig,1998). This is markedly different from Bacteria, which areunified by the universal presence of murein in their cellwall (with the exception of a few derived lineages that lostthis feature). The ubiquity of murein in bacterial cell enve-lopes suggests that it was an early invention in this lineage(Kandler and Konig, 1998), which could have played animportant role in maintaining its cohesion. The switch inmembrane lipid stereospecificity could have been a com-parable event for Archaea, isolating proto-archaea fromother prokaryotes and allowing the differentiation of thefirst members of this domain of life.
Much has yet to be discovered concerning the geneticsof isoprenoid lipid biosynthesis in archaea. Key enzymeslike the archaeal analogues catalysing the last two stepsof the mevalonate pathway, the proteins responsible forthe hydrogenation of the isoprenoid side-chains and theenzymes joining diether lipids to form cyclic tetraetherlipids, are still uncharacterized. However, the informationcurrently available about genes involved in biosynthesis
of isoprenoid lipids tell us that this apparatus evolvedthrough the co-option of ancestral enzymes for novel func-tions (GGGP synthase, G1P dehydrogenase), tinkeringwith specificity (GGPP/FGPP synthases, GGGP/FGGPsynthases), orthologous displacement (HMGR), inventionof archaeal specific analogues (archaeal analogues ofPMK and PPMD), integration of components fromeukaryotes and bacteria (PMK, PPMD and IDI1), rapiddivergence (GGGP synthase) and LGT within (IDI2 inHalobacteriales) and between (GGGP synthase) archaealorders.
Experimental procedures
Archaeal strains and DNA extraction
The following strains of extremely halophilic archaea wereused in this study: Halobacterium salinarum JCM9120, Halo-bacterium salinarum ATCC19700, Halococcus morrhuaeNRC16008, Haloferax denitrificans ATCC35960, Haloferaxmediterranei ATCC33500, Halorhabdus utahensis DSM12940, Halorubrum distributum JCM9100, Haloterrigenaturkmenica VKM B-1734, Natrialba asiatica 172P1 JCM9576,Natrinema versiforme XF10 JCM10478, Natronobacteriumgregoryi NCMB2189, Natronobacterium sp. SSL-6 (a giftfrom Dr V. Upasani), Natronomonas pharaonis DSM2160,Natronorubrum sp. Tenzan-10 JCM10938 and Natrinema sp.XA3-1 (a gift from Dr P. Zhou). Genomic DNA was extractedfrom these strains using the protocol from Wilson (1994).
Polymerase chain reaction (PCR) amplification, cloning and sequencing
The amplification of PPMD, IDI1, IDI2, GGGPS and GDgenes was done in two steps. First, several degenerate prim-ers were designed from available database sequences for theamplification of each gene from the genomic DNA of allavailable strains of extremely halophilic archaea. Sequencesobtained for each gene were subsequently aligned to designbetter degenerate primers for all five genes: PPMD_forward1:5¢-CTCGTGAARTAYCAYGGSATG-3¢, PPMD_reverse1 5¢-GGCTTCCAGTASACCCNCC-3¢, IDI1_forward1 5¢-TGGGACACCTNCTGGGAYGG-3¢, IDI1_reverse1 5¢-GCGATCTCGAACCANGGRCA-3¢, IDI2_foward1 5¢-ATCGACTCAATGACNGGN GG-3¢, IDI2_reverse1 5¢-GT CTACGTCACCYTCNGGYTG-3¢, GGGPS_foward1 5¢-GTGCCCCTCTAYCAGGARCC-3¢, GGGPS_reverse1 5¢-CTGGATGCCRCCRCCRTAGAA-3¢, GGGPS_foward2 5¢-CACGTCACGAARTGNGAYCC-3¢, GGGPS_reverse2 5¢-CCCACCACGACSGCGTCSGC-3¢,GD_foward1 5¢-CCGTCCACGTAKGTNCARGG-3¢, GD_reverse1 5¢-ATATTGACCTTYTCNCCRTG-3¢, GD_foward25¢-CTGGCGACNTCCTTYGARGC-3¢, GD_reverse2 5¢-GGCTCATCGTGDATNGTYTC-3¢. Each gene was amplified fromtwo independent PCR reactions. Amplifications were carriedout in a final volume of 25 ml containing 1–5 ng of templateDNA, 1¥ PCR buffer, 2.5 mM MgCl2, 0.2 mM dNTPs, 1.0 mMof each primer, and 0.5–1 U of Platinum Taq High FidelityDNA polymerase (Invitrogen). The reactions were performedwith an initial denaturation at 94∞C for 1 min, 30 cycles with

526 Y. Boucher, M. Kamekura and W. F. Doolittle
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 515–527
a denaturation at 94∞C for 30 s., primer annealing at 48–52∞Cfor 30 s., and primer extension at 72∞C for 1 min. The PCRproducts were gel purified with the MinElute kit (Qiagen) andcloned in TopoTA (Invitrogen). Two clones were sequencedfrom both strands for each PCR product using a LiCor4000 L automated sequencer. The sequences determined inthis study have been submitted to the EMBL nucleotidesequence database and assigned accession numbersAJ566212 and AJ564466 to AJ564495.
Similarity searches, multiple sequence alignment and phylogenetic analysis
All genes sequences were retrieved from the NCBI website(http://www.ncbi.nlm.nih.gov/) through similarity searchesperformed using BLASTP (http://www.ncbi.nlm.nih.gov/BLAST/).A biochemically characterized archaeal homologue of eachenzyme was used as the query. Functional homology ofsignificant hits (e-value inferior to 1 ¥ 10-5) was inferred ifamino acid motifs important to the functionality of the partic-ular enzyme were conserved. Preliminary sequence datafrom the unfinished genomes of Haloferax volcanii(http://wit-scranton.mbi.scranton.edu/Haloferax/) and Haloar-cula marismortui (http://zdna2.umbi.umd.edu/cgi-bin/BLAST/BLAST.pl) were obtained from their respective websites. Theretrieved amino acid sequences and the new sequences fromthis study were aligned using CLUSTALW (Thompson et al.,1994). The alignment was subsequently edited manually toremove ambiguous characters and regions corresponding tothe primers in novel sequences. 5¢-and 3¢ regions absentfrom the novel sequences (that are only partial) were kept inthe alignment if phylogenetically informative. The number ofsites used in the different protein sequence alignments wereas follows (the first number is the length of the novel partialsequences and the second number is the full length of thealignment): MVK/PMK (165\165), PPMD (117/204), IDI1 (87/139), IDI2 (78/243), GGPPS (198/198) GGGPS (131/177),GD/G1PD (146/268). Maximum likelihood phylogenetic anal-yses were performed using PROML with the JTT amino acidsubstitution matrix, a rate heterogeneity model with gamma-distributed rates over four categories with the a parameterestimated using TREE-PUZZLE, global rearrangements andrandomized input order of sequences (10 jumbles). Bootstrapsupport values represent a consensus (obtained usingCONSENSE) of 100 Fitch-Margoliash distance trees (obtainedusing PUZZLEBOOT and FITCH) from pseudo-replicates(obtained using SEQBOOT) of the original alignment. The set-tings of PUZZLEBOOT were the same as those used for PROML,except that no global rearrangements and randomized inputorder of sequences are available in this program. PROML,CONSENSE, FITCH and SEQBOOT are from the PHYLIP packageversion 3.6a (http://evolution.genetics.washington.edu/phylip.html). TREE-PUZZLE and PUZZLEBLOOT can be obtainedfrom the programs website (http://www.tree-puzzle.de).
Phylogenetic analyses were also carried at the DNA levelfor the taxa belonging to the order Halobacteriales. Theseanalyses were performed with PAUP* 4.04b (Swofford, 1998)applying the heuristic-search option and using the TBRbranch-swapping algorithm. Maximum likelihood was usedas the tree reconstruction method, with the nucleotide sub-stitution model, gamma rates parameter a, proportion of
invariable sites and nucleotide frequencies determinedindependently for each gene using MODELTEST (Posada andCrandall, 1998). The confidence of each node wasdetermined by building a consensus tree of 100 bootstrappseudo-replicates.
Acknowledgements
The authors thank Christophe Douady for technical com-ments and suggestions on phylogenetic analysis. We arealso grateful to Dr V. Upasani for providing us with theNatronobacterium sp. SSL-6 strain and to Dr P. Zhou for theNatrinema sp. XA3-1 strain.
References
Boucher, Y., and Doolittle, W.F. (2000) The role of lateralgene transfer in the evolution of isoprenoid biosynthesispathways. Mol Microbiol 37: 703–716.
Boucher, Y., Huber, H., L’Haridon, S., Stetter, K.O., andDoolittle, W.F. (2001) Bacterial origin for the isoprenoidbiosynthesis enzyme HMG-CoA reductase of the archaealorders Thermoplasmatales and Archaeoglobales. Mol BiolEvol 18: 1378–1388.
Chen, A., and Poulter, C.D. (1993) Purification and charac-terization of farnesyl diphosphate/geranylgeranyl diphos-phate synthase. A thermostable bifunctional enzyme fromMethanobacterium thermoautotrophicum. J Biol Chem268: 11002–11007.
De Rosa, M., and Gambacorta, A. (1988) The lipids ofarchaebacteria. Prog Lipid Res 27: 153–175.
Dennis, P.P., and Shimmin, L.C. (1997) Evolutionarydivergence and salinity-mediated selection in halophilicarchaea. Microbiol Mol Biol Rev 61: 90–104.
Iordanescu, S. (1993) Characterization of the Staphylococ-cus aureus chromosomal gene pcrA, identified by muta-tions affecting plasmid pT181 replication. Mol Gen Genet241: 185–192.
Kamekura, M., and Kates, M. (1999) Structural diversity ofmembrane lipids in members of Halobacteriaceae. BiosciBiotechnol Biochem 63: 969–972.
Kandler, O., and Konig, H. (1998) Cell wall polymers inArchaea. Cellular Mol Life Sci 54: 305–308.
Kaneda, K., Kuzuyama, T., Takagi, M., Hayakawa, Y., andSeto, H. (2001) An unusual isopentenyl diphosphateisomerase found in the mevalonate pathway gene clusterfrom Streptomyces sp. strain CL190. Proc Natl Acad SciUSA 98: 932–937.
Kates, M., and Kushwaha, N. (1978) Biochemistry of thelipids of extremely halophilic bacteria. In Energetics andStructure of Halophilic Microorganisms. Caplan, S.R.,and Ginzburg, M., (eds). Amsterdam: Elsevier, pp. 461–480.
Kellogg, B.A., and Poulter, C.D. (1997) Chain elongation inthe isoprenoid biosynthetic pathway. Curr Opin Chem Biol1: 570–578.
Lange, B.M., Rujan, T., Martin, W., and Croteau, R. (2000)Isoprenoid biosynthesis: the evolution of two ancient anddistinct pathways across genomes. Proc Natl Acad SciUSA 97: 13172–13177.

Isoprenoid lipid biosynthesis in archaea 527
© 2004 Blackwell Publishing Ltd, Molecular Microbiology, 52, 515–527
Matte-Tailliez, O., Brochier, C., Forterre, P., and Philippe, H.(2002) Archaeal phylogeny based on ribosomal proteins.Mol Biol Evol 19: 631–639.
Morii, H., Nishihara, M., and Koga, Y. (2000) CTP: 2,3-di-O-geranylgeranyl-sn-glycero-1-phosphate cytidyltransferasein the methanogenic archaeon Methanothermobacter ther-moautotrophicus. J Biol Chem 275: 36568–36574.
Ng, W.V., Kennedy, S.P., Mahairas, G.G., Berquist, B., Pan,M., Shukla, H.D., et al. (2000) Genome sequence of Halo-bacterium species NRC-1. Proc Natl Acad Sci USA 97:12176–12181.
Nishihara, M., and Koga, Y. (1995) sn-glycerol-1-phosphatedehydrogenase in Methanobacterium thermoautotrophi-cum: key enzyme in biosynthesis of the enantiomericglycerophosphate backbone of ether phospholipids ofarchaebacteria. J Biochem (Tokyo) 117: 933–935.
Nishihara, M., Yamazaki, T., Oshima, T., and Koga, Y. (1999)sn-glycerol-1-phosphate-forming activities in Archaea:separation of archaeal phospholipid biosynthesis and glyc-erol catabolism by glycerophosphate enantiomers. J Bac-teriol 181: 1330–1333.
Ohnuma, S., Hirooka, K., Hemmi, H., Ishida, C., Ohto, C.,and Nishino, T. (1996) Conversion of product specificityof archaebacterial geranylgeranyl-diphosphate synthase.Identification of essential amino acid residues for chainlength determination of prenyltransferase reaction. J BiolChem 271: 18831–18837.
Ohnuma, S., Hirooka, K., Ohto, C., and Nishino, T. (1997)Conversion from archaeal geranylgeranyl diphosphatesynthase to farnesyl diphosphate synthase. Two aminoacids before the first aspartate-rich motif solely determineeukaryotic farnesyl diphosphate synthase activity. J BiolChem 272: 5192–5198.
Ohnuma, S., Hirooka, K., Tsuruoka, N., Yano, M., Ohto, C.,Nakane, H., and Nishino, T. (1998) A pathway wherepolyprenyl diphosphate elongates in prenyltransferase.Insight into a common mechanism of chain length deter-mination of prenyltransferases. J Biol Chem 273: 26705–26713.
Posada, D., and Crandall, K.A. (1998) MODELTEST: testing
the model of DNA substitution. Bioinformatics 14: 817–818.
Shimada, H., Nemoto, N., Shida, Y., Oshima, T., and Yam-agishi, A. (2002) Complete polar lipid composition ofThermoplasma acidophilum HO-62 determined by high-performance liquid chromatography with evaporative light-scattering detection. J Bacteriol 184: 556–563.
Smit, A., and Mushegian, A. (2000) Biosynthesis of iso-prenoids via mevalonate in Archaea: the lost pathway.Genome Res 10: 1468–1484.
Soderberg, T., Chen, A., and Poulter, C.D. (2001) Gera-nylgeranylglyceryl phosphate synthase. Characterizationof the recombinant enzyme from Methanobacterium ther-moautotrophicum. Biochemistry 40: 14847–14854.
Swofford, D.L. (1998) PAUP*. Phylogenetic Analysis UsingParsimony (* and Other Methods). Sunderland, Massa-chusets: Sinauer Associates.
Tachibana, A. (1994) A novel prenyltransferase, farnesylger-anyl diphosphate synthase, from the haloalkaliphilicarchaeon, Natronobacterium pharaonis. FEBS Lett 341:291–294.
Tachibana, A., Yano, Y., Otani, S., Nomura, N., Sako, Y.,and Taniguchi, M. (2000) Novel prenyltransferase geneencoding farnesylgeranyl diphosphate synthase from ahyperthermophilic archaeon, Aeropyrum pernix. Molecu-larevolution with alteration in product specificity. Eur JBiochem 267: 321–328.
Thompson, J.D., Higgins, D.G., and Gibson, T.J. (1994)CLUSTAL W: improving the sensitivity of progressive multiplesequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. NucleicAcids Res 22: 4673–4680.
Wilson, K. (1994) Preparation of genomic DNA from bacteria.In Current Protocols in Molecular Biology Vol. 1. Ausabel,F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidmon,J.A., Stuhl, K., and Smith, J.A. (eds). New York: John Wileyand Sons, pp. 2.4.1.–2.4.5.
Zhang, D., and Poulter, C.D. (1993) Biosynthesis of archae-bacterial ether lipids. Formation of ether linkages by pre-nyltransferases. J Am Chem Soc 115: 1270–1277.
Related Documents











