Mind the gaps: Theoretical and empirical Mind the gaps: Theoretical and empirical aspects of plant ecology and population aspects of plant ecology and population genetics genetics S. peruvianu m S. chilense 08.03.2013 Aurélien Tellier Populationsgenetik TUM Weihenstephan, Freising

Mind the gaps: Theoretical and empirical aspects of plant ecology and population genetics S. peruvianum S. chilense 08.03.2013 Aurélien Tellier Populationsgenetik.
Dec 26, 2015
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mind the gaps: Theoretical and empirical Mind the gaps: Theoretical and empirical
aspects of plant ecology and population aspects of plant ecology and population
geneticsgenetics
S. peruvianum
S. chilense
08.03.2013
Aurélien TellierPopulationsgenetik
TUM Weihenstephan, Freising
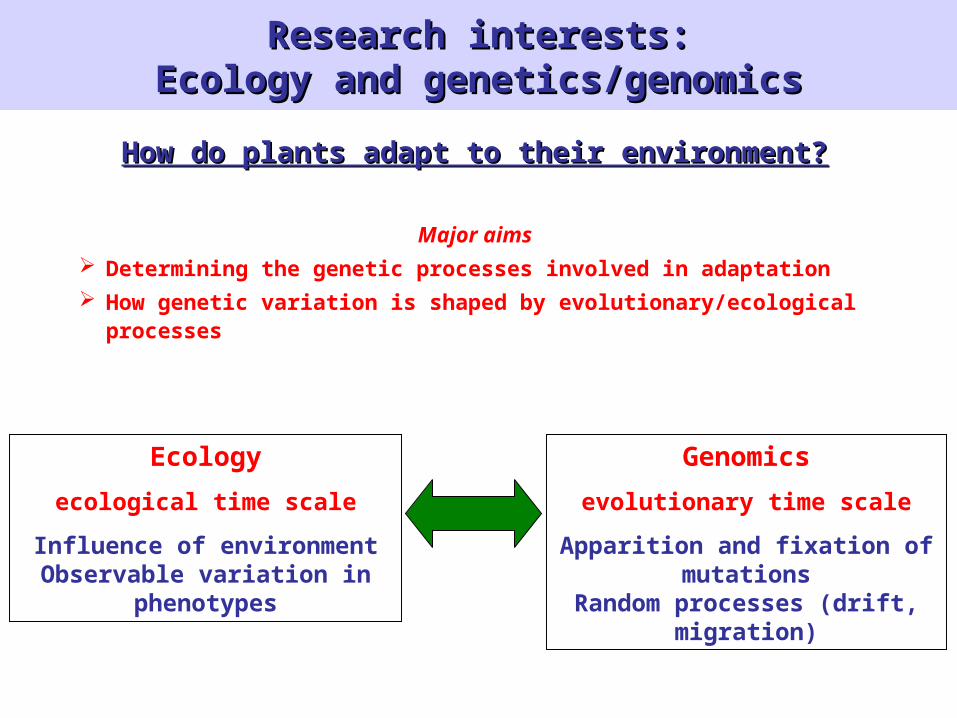
Research interests:Research interests:Ecology and genetics/genomicsEcology and genetics/genomics
How do plants adapt to their environment?How do plants adapt to their environment?
Major aims Determining the genetic processes involved in adaptation How genetic variation is shaped by evolutionary/ecological
processes
Ecology
ecological time scale
Influence of environmentObservable variation in
phenotypes
Genomics
evolutionary time scale
Apparition and fixation of mutations
Random processes (drift, migration)

Research interests: Two systemsResearch interests: Two systems
Plant-parasite coevolution
Seed banks and seed dormancy
2011
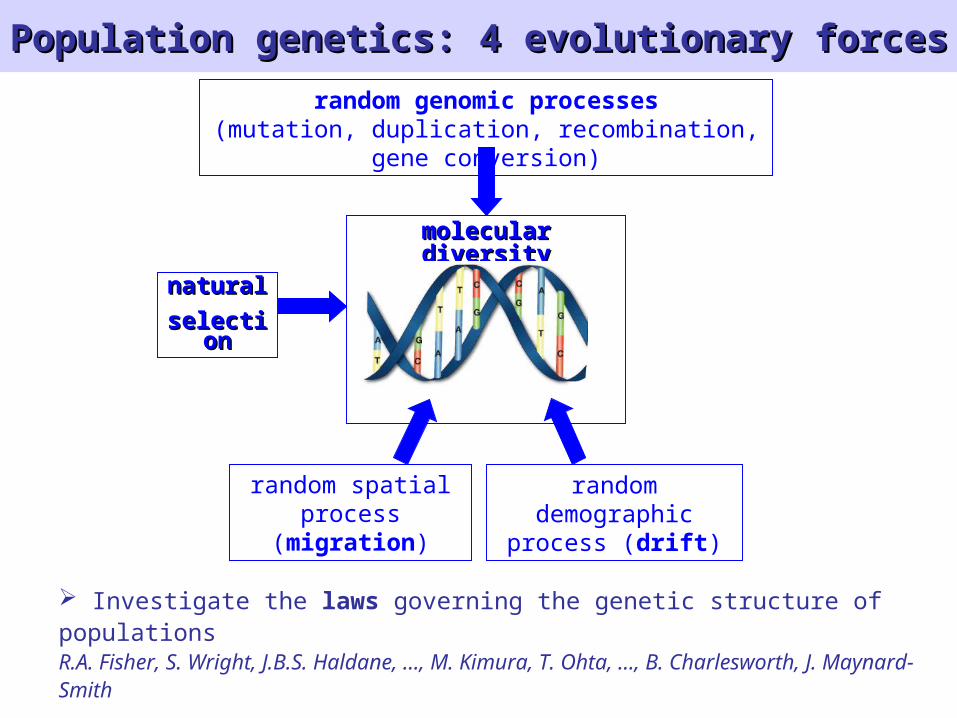
Population genetics: 4 evolutionary forcesPopulation genetics: 4 evolutionary forces
random genomic processes(mutation, duplication, recombination, gene
conversion)
naturalnatural
selectiselectionon
random demographic
process (drift)
random spatialprocess
(migration)
molecular diversitymolecular diversity
Investigate the laws governing the genetic structure of populationsR.A. Fisher, S. Wright, J.B.S. Haldane, …, M. Kimura, T. Ohta, …, B. Charlesworth, J. Maynard-Smith
Coalescent theory is a tool to infer past evolution from sequence data
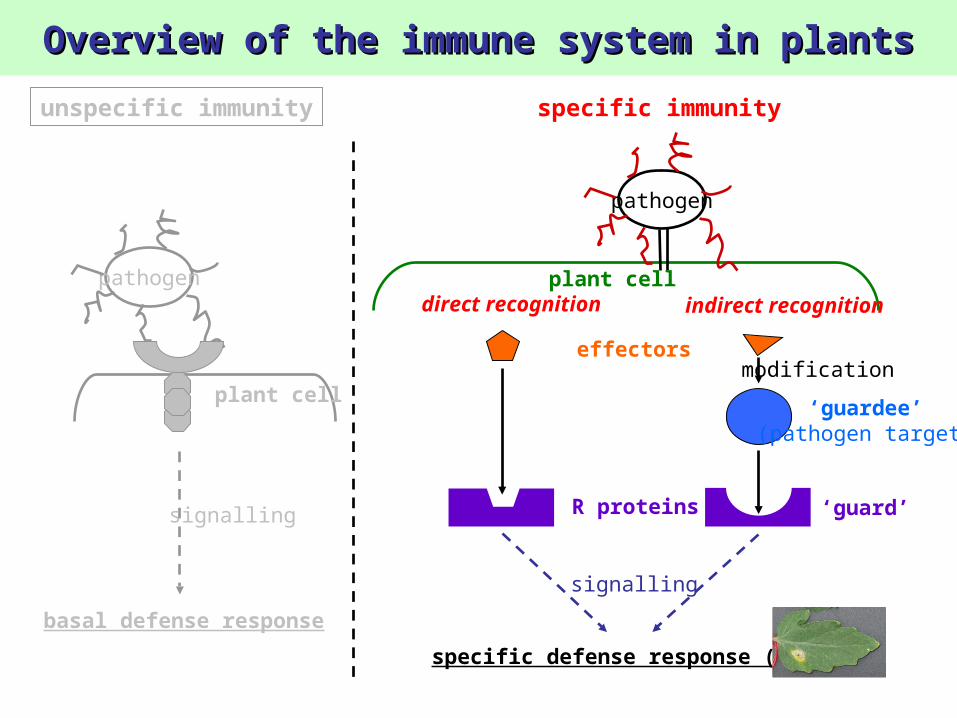
pathogen
plant cell
pathogen
effectors
direct recognition indirect recognitionplant cell
R proteins
modification
‘guardee’(pathogen target)
‘guard’
specific defense response (HR)
signalling
signalling
basal defense response
unspecific immunity specific immunity
Overview of the immune system in plantsOverview of the immune system in plants
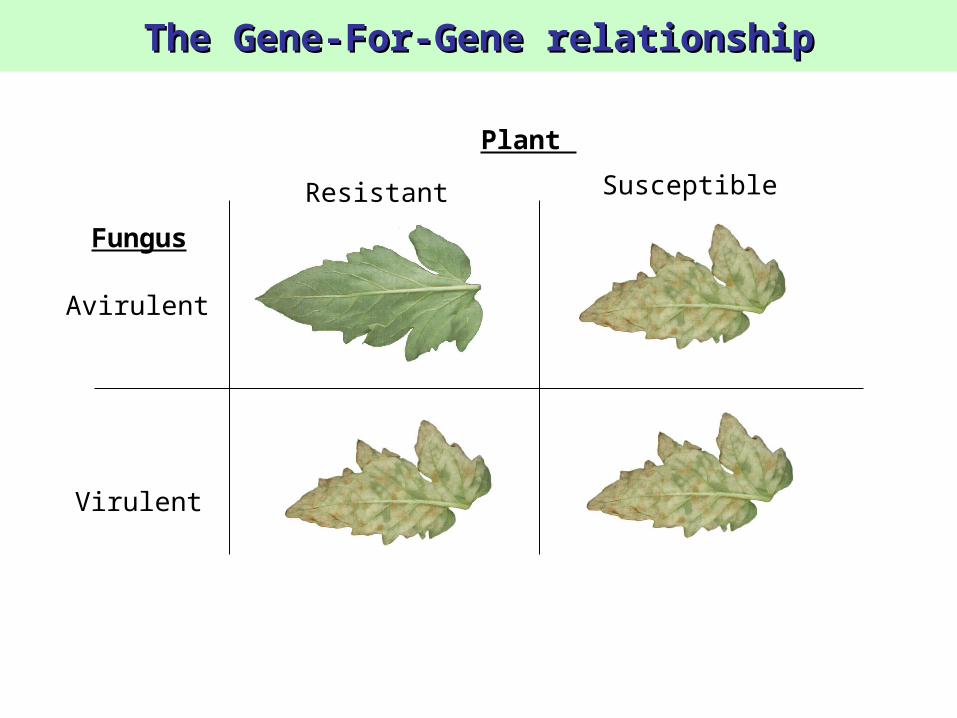
The Gene-For-Gene relationshipThe Gene-For-Gene relationship
Plant
Resistant Susceptible
Fungus
Avirulent
Virulent
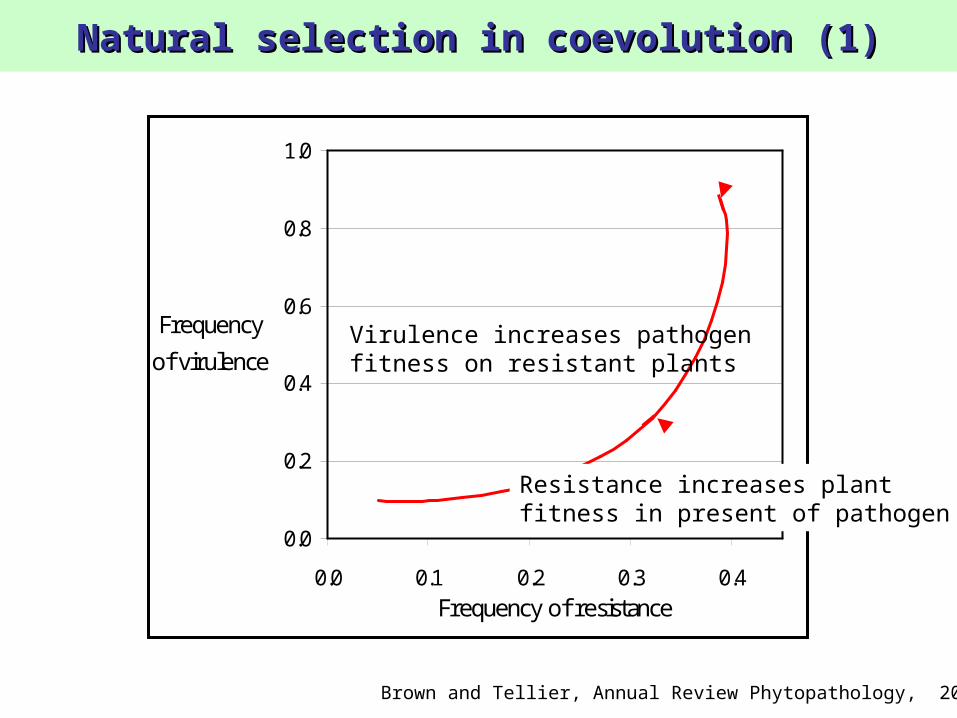
Natural selection in coevolution (1)Natural selection in coevolution (1)
0.0
0.2
0.4
0.6
0.8
1.0
0.0 0.1 0.2 0.3 0.4Frequency of resistance
Frequency
of virulence
0.0
0.2
0.4
0.6
0.8
1.0
0.0 0.1 0.2 0.3 0.4Frequency of resistance
Frequency
of virulence
Resistance increases plantfitness in present of pathogen
Virulence increases pathogenfitness on resistant plants
Brown and Tellier, Annual Review Phytopathology, 2011
0.0
0.2
0.4
0.6
0.8
1.0
0.0 0.1 0.2 0.3 0.4Frequency of resistance
Frequency
of virulence
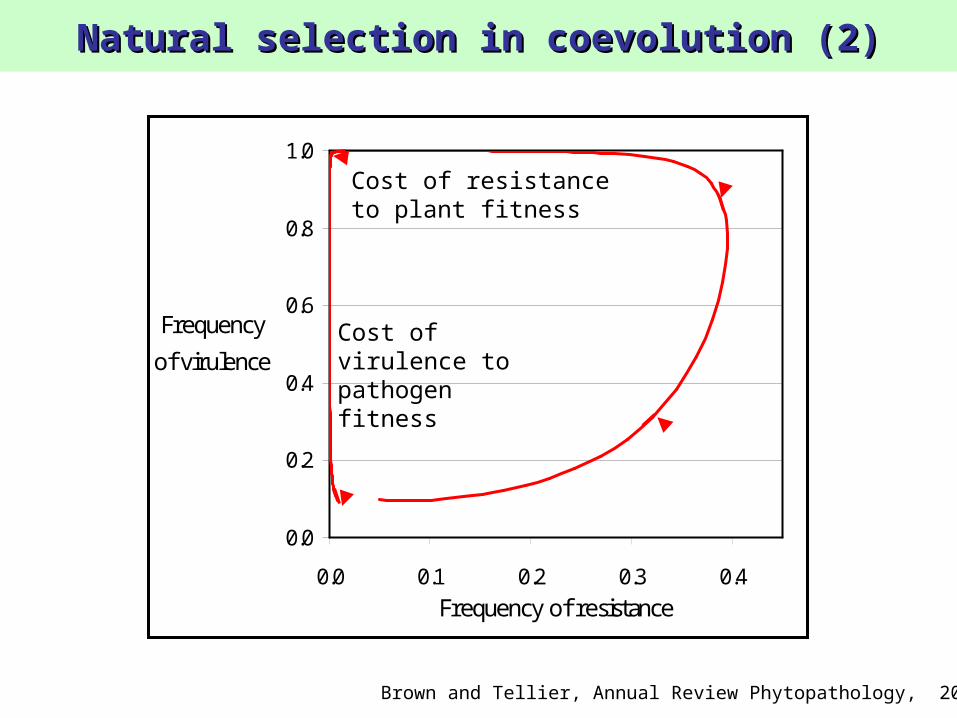
Natural selection in coevolution (2)Natural selection in coevolution (2)
0.0
0.2
0.4
0.6
0.8
1.0
0.0 0.1 0.2 0.3 0.4Frequency of resistance
Frequency
of virulence
0.0
0.2
0.4
0.6
0.8
1.0
0.0 0.1 0.2 0.3 0.4Frequency of resistance
Frequency
of virulence
Cost of resistanceto plant fitness
Cost of virulence to pathogen fitness
Brown and Tellier, Annual Review Phytopathology, 2011
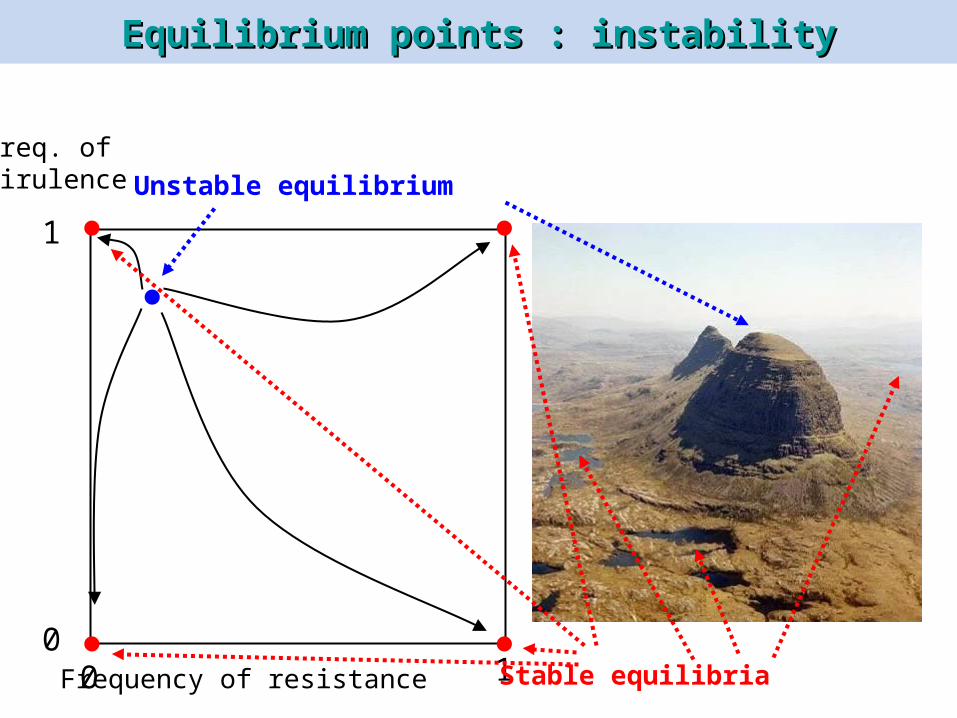
Equilibrium points : instabilityEquilibrium points : instability
Frequency of resistance
1
100
Freq. ofvirulence Unstable equilibrium
Stable equilibria
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
0,00 0,20 0,40 0,60 0,80
RES frequency
av
r frequency
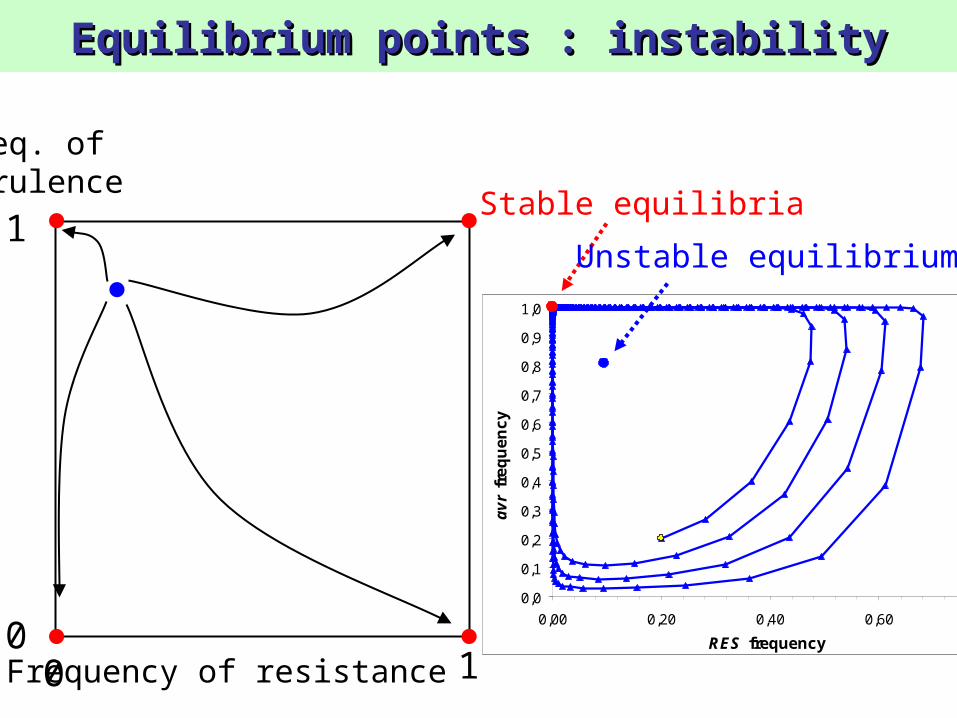
Frequency of resistance
1
100
Freq. ofvirulence
Stable equilibria
Unstable equilibrium
Equilibrium points : instabilityEquilibrium points : instability
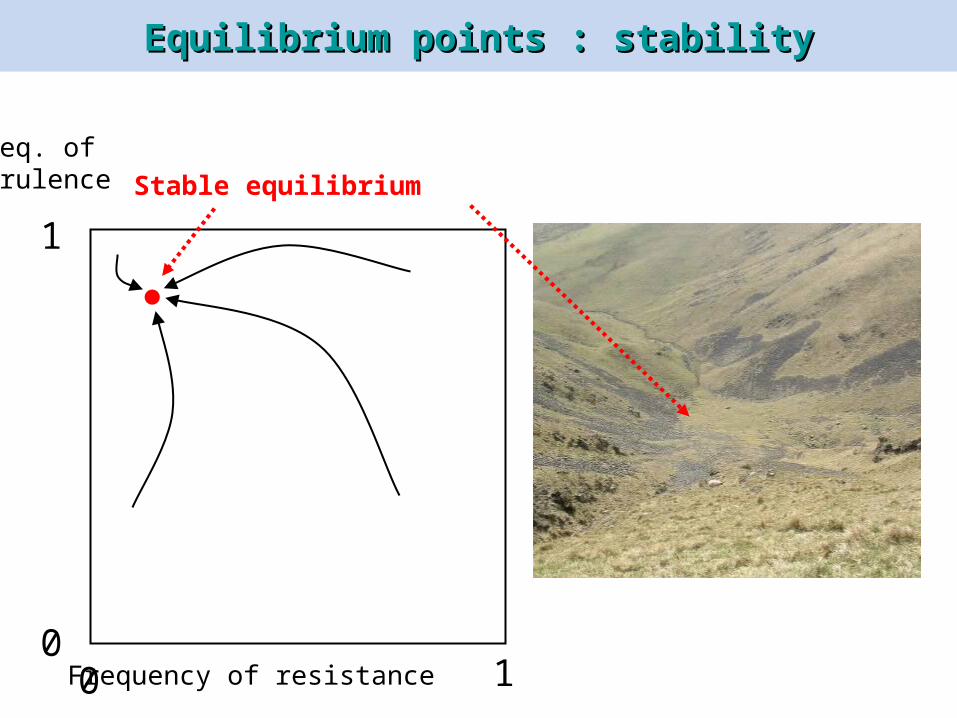
Equilibrium points : stabilityEquilibrium points : stability
Frequency of resistance
1
100
Freq. ofvirulence Stable equilibrium
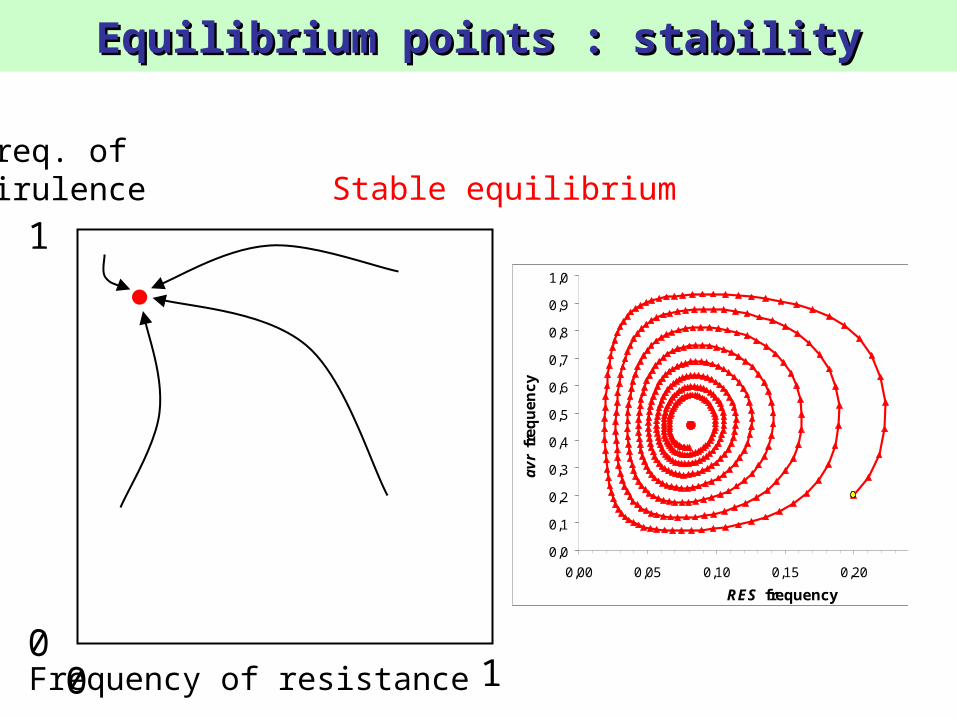
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
0,00 0,05 0,10 0,15 0,20 0,25 0,30
RES frequency
av
r frequency
Frequency of resistance
1
100
Freq. ofvirulence Stable equilibrium
Equilibrium points : stabilityEquilibrium points : stability

Why the simple model is unstable (1)Why the simple model is unstable (1)
Taking log of ratio, ρ = log(R/r) … maths easier!
The change in R depends on A but not on R itself
=R( r ) ′ R
r.(1−u)(1−s+sA)
1−ss = cost of diseaseu = cost of resistance
log ρ′ − log ρ = Δρ log ρ′ − log ρ = log(1−u) + log(1−s−sA) − log(1−s)

Taking log of ratio, α = log(A/a)log α′ − log α = Δα log ρ′ − log ρ = log(1−cR) − log(1−b)
The change in A depends on R but not on A itself
=A( a ) ′ A
a. 1−cR
1−bc = cost of AVR on RES
plantb = cost of virulence
Hence dΔρ/dR + dΔα/dA = 0 and stability is impossible
Why the simple model is unstable (1)Why the simple model is unstable (1)

Requirements for stable polymorphismRequirements for stable polymorphism
Tellier and Brown, Proc. Roy. Soc. B, 2007
/ /
.// /
/
/
/R r
RA
R r R r
A aA
R r
A aa
Ra
A
A
The eigenvalues 1,2 = i of the Jacobian Matrix should be in a circle centered on (-1,0) in the complex plane
Jacobian Matrix
(-1,0)
-
/ /R r A a
R A
<0 the sum of the diagonal coefficients of
the Jacobian matrix

Requirements for stable polymorphismRequirements for stable polymorphism
Avirulence selects for increased resistance
Resistance selects for increased virulence
= indirect frequency-dependent selection
→ graph of alleles frequencies (R,a) spirals
Selection for RES weaker as RES more common
Or selection for avr weaker as avr more common
= direct frequency-dependent selection
→ graph spirals to a stable equilibrium point
Tellier and Brown, Proc. Roy. Soc. B, 2007

Boom and Bust cycle: stable and unstable Boom and Bust cycle: stable and unstable dynamicsdynamics
Brown and Tellier Ann Rev Phytopat 2011
Factors promoting stable dynamics:
Biological factors (life history traits):
Polycylicity = several pathogen generations/host generation (Tellier
and Brown, Proc Roy Soc B, 2007)
Perenniality or seed banks in host (Tellier and Brown, Am Nat, 2009)
Genetic factors: Epistasis among R-genes and virulence genes (Tellier and
Brown, Genetics, 2007)
Ecological factors: Environmental variability in space (Laine and Tellier
Oikos 2008; Tellier and Brown BMC Evol Biol 2011)

Stable and unstable dynamicsStable and unstable dynamics
Brown and Tellier Ann Rev Phytopat 2011
Biological factors (life history traits): Polycylicity, perenniality or seed
banks
Genetic factors: Epistasis
Ecological factors: Environmental variability in space (see work by Laine,
Thrall, Burdon,…)
Expect to find often stable dynamics in natural populations!
(and conversely more unstable dynamics in agro-systems)
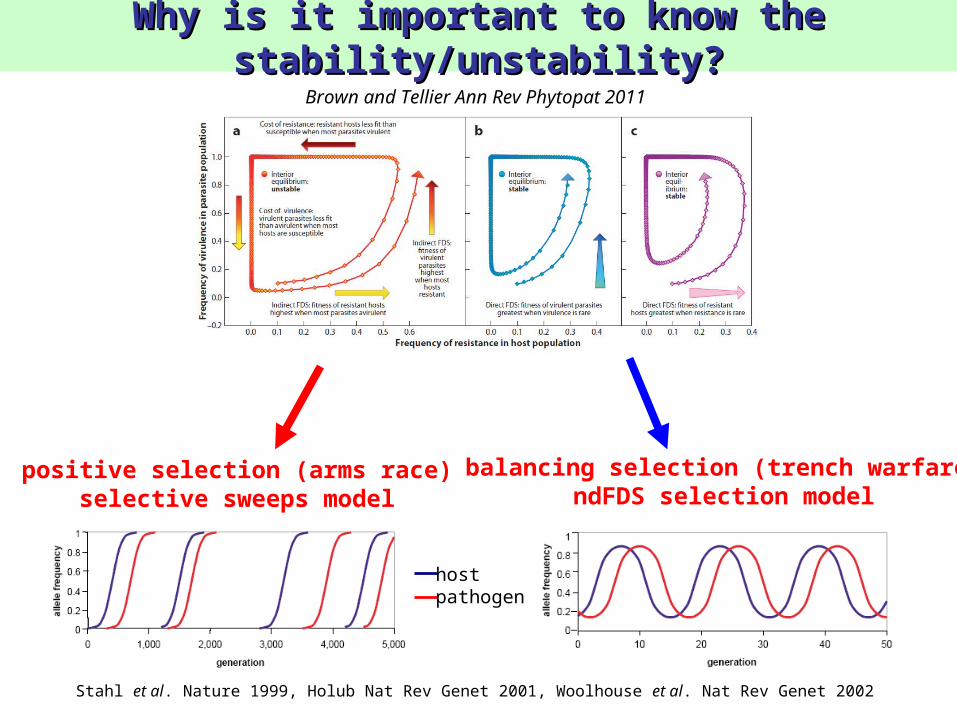
Why is it important to know the Why is it important to know the stability/unstability?stability/unstability?
hostpathogen
Stahl et al. Nature 1999, Holub Nat Rev Genet 2001, Woolhouse et al. Nat Rev Genet 2002
balancing selection (trench warfare)ndFDS selection model
positive selection (arms race)selective sweeps model
Brown and Tellier Ann Rev Phytopat 2011
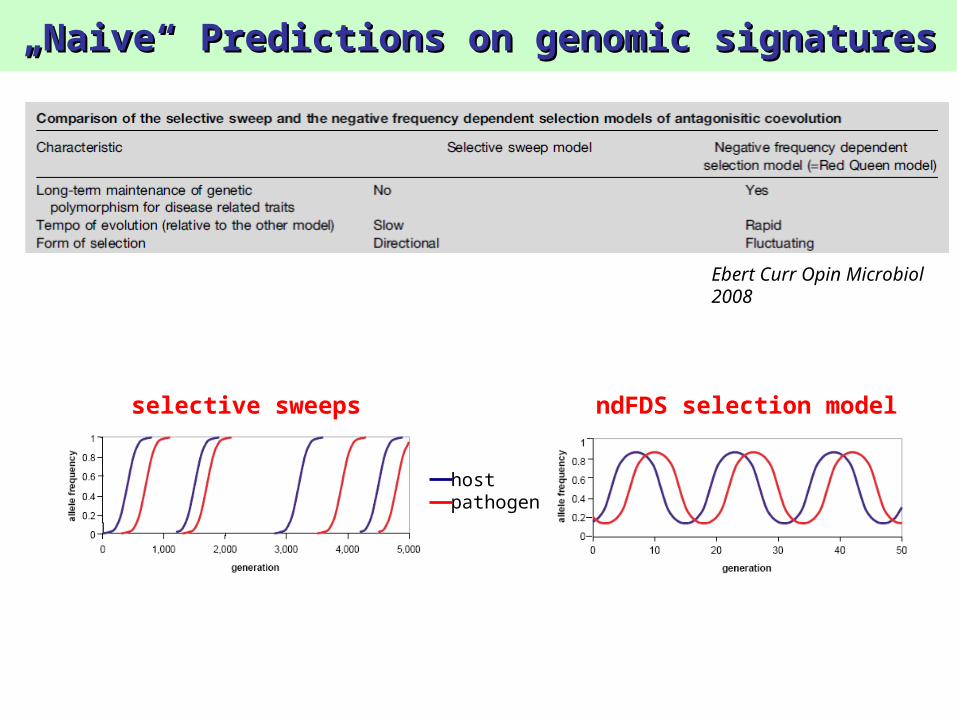
„„Naive“ Predictions on genomic signaturesNaive“ Predictions on genomic signatures
Ebert Curr Opin Microbiol 2008
hostpathogen
ndFDS selection modelselective sweeps
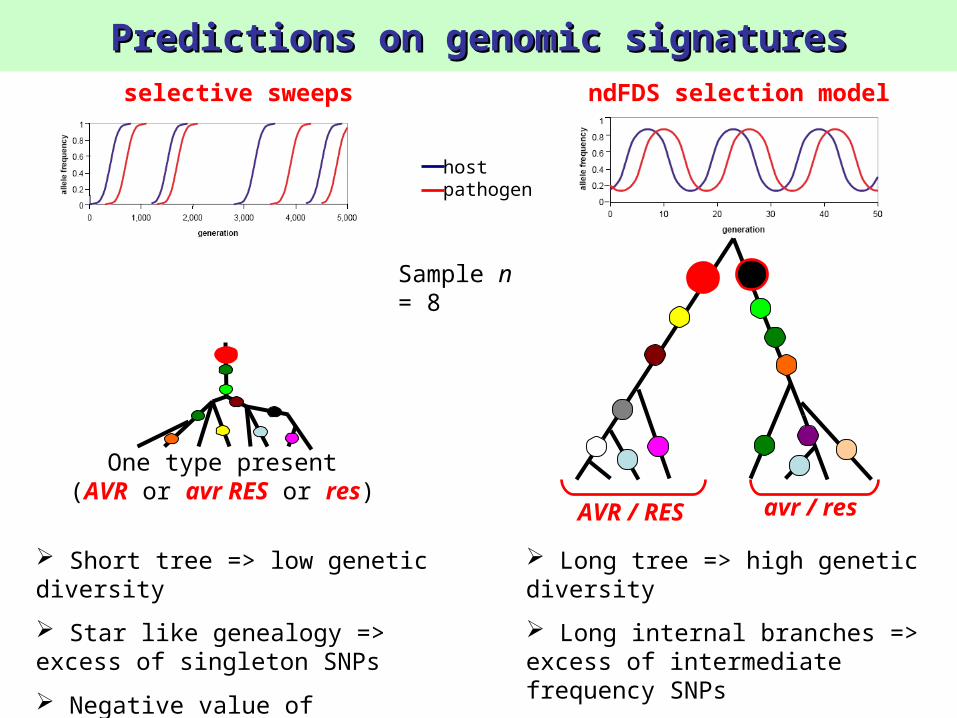
Predictions on genomic signaturesPredictions on genomic signatures
hostpathogen
ndFDS selection modelselective sweeps
Short tree => low genetic diversity
Star like genealogy => excess of singleton SNPs
Negative value of Tajima’s D (<0)
Long tree => high genetic diversity
Long internal branches => excess of intermediate frequency SNPs
Positive value of Tajima’s D (>0)
Sample n = 8
One type present(AVR or avr RES or res)
AVR / RES avr / res
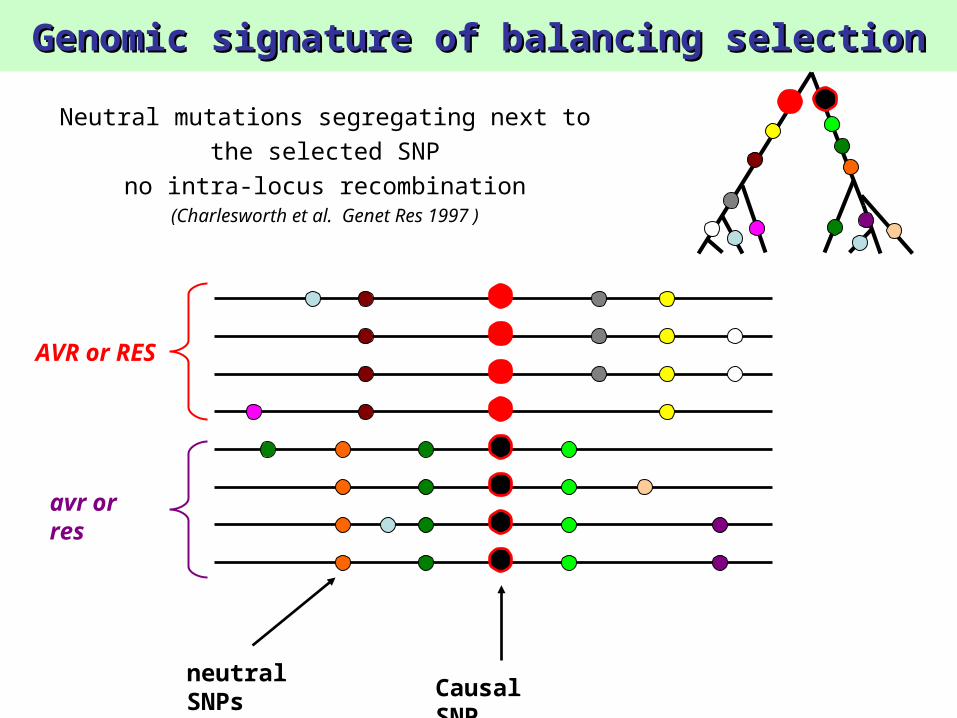
Genomic signature of balancing selectionGenomic signature of balancing selection
Neutral mutations segregating next to the selected SNP
no intra-locus recombination(Charlesworth et al. Genet Res 1997 )
AVR or RES
avr or res
Causal SNP
neutral SNPs
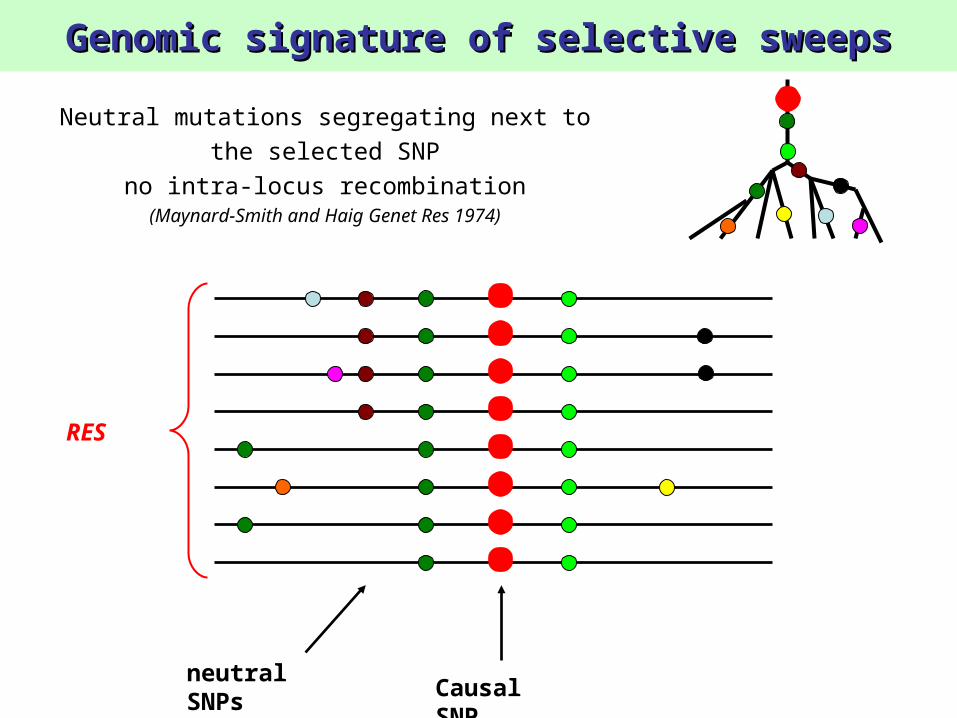
Genomic signature of selective sweepsGenomic signature of selective sweeps
RES
Causal SNP
neutral SNPs
Neutral mutations segregating next to the selected SNP
no intra-locus recombination(Maynard-Smith and Haig Genet Res 1974)
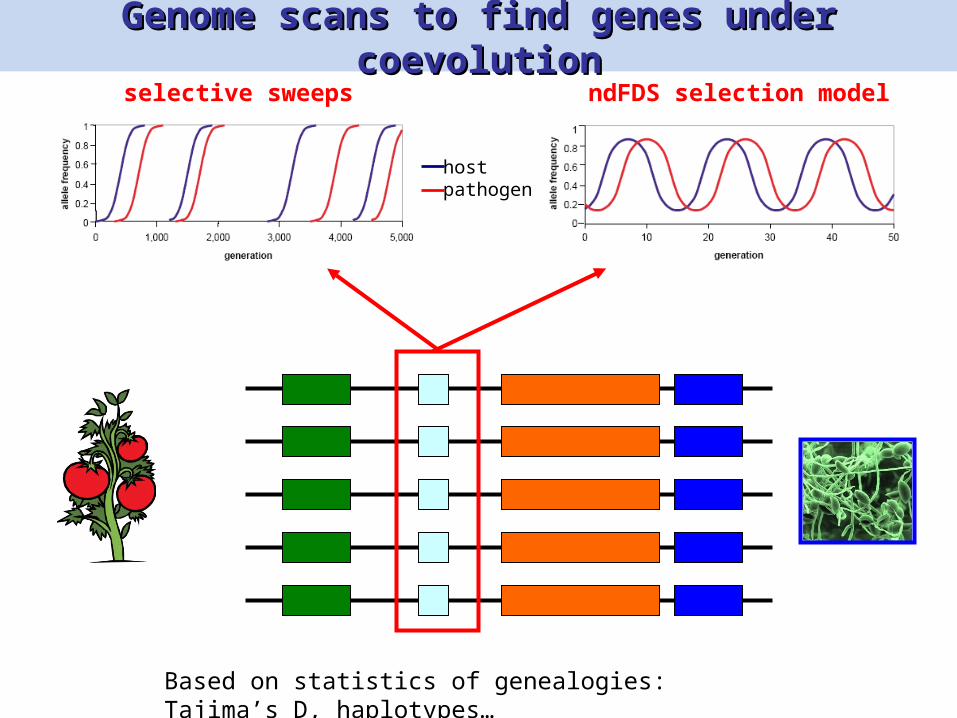
Genome scans to find genes under Genome scans to find genes under coevolutioncoevolution
hostpathogen
ndFDS selection modelselective sweeps
Based on statistics of genealogies: Tajima’s D, haplotypes…
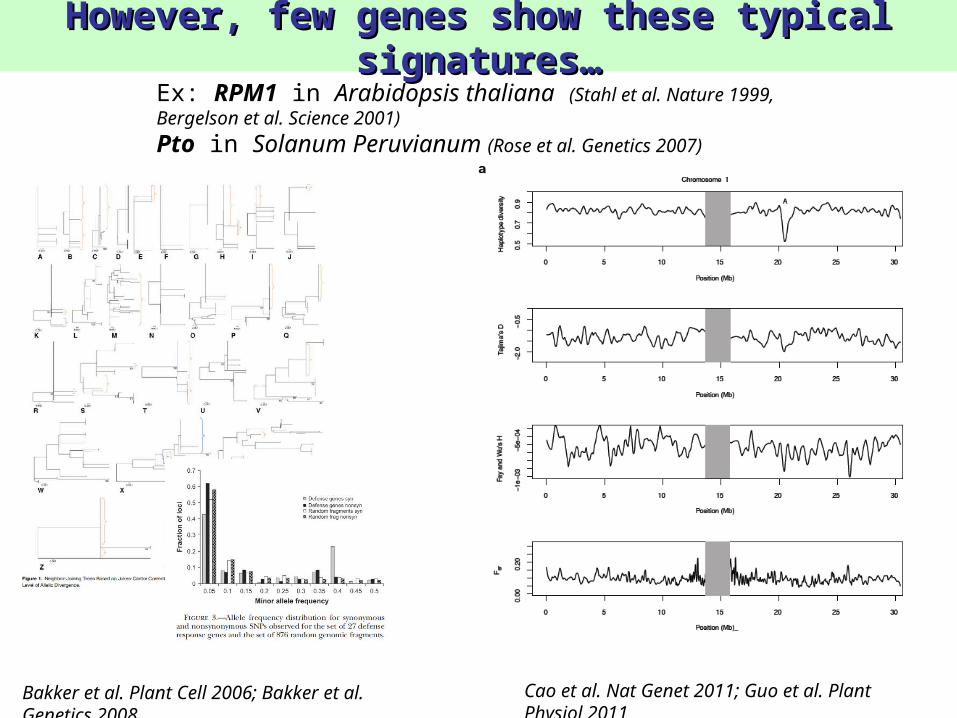
However, few genes show these typical However, few genes show these typical signatures…signatures…
Bakker et al. Plant Cell 2006; Bakker et al. Genetics 2008
Cao et al. Nat Genet 2011; Guo et al. Plant Physiol 2011
Ex: RPM1 in Arabidopsis thaliana (Stahl et al. Nature 1999, Bergelson et al. Science 2001)
Pto in Solanum Peruvianum (Rose et al. Genetics 2007)

A major draw back of current theory is that it is based on deterministic models and short-term ecological models
Questions:
1) What is the role of genetic drift for stability or allele fixation?
2) In finite populations, can stable dynamics lead to fixation of alleles? Under which conditions?
3) Are dynamics in the arms race model slower than in the trench warfare model?
4) Expectations for genomic signatures under GFG models?
Why so little genomic evidence for Why so little genomic evidence for coevolution?coevolution?
Exemple of a stable model in finite population (N = 3000)
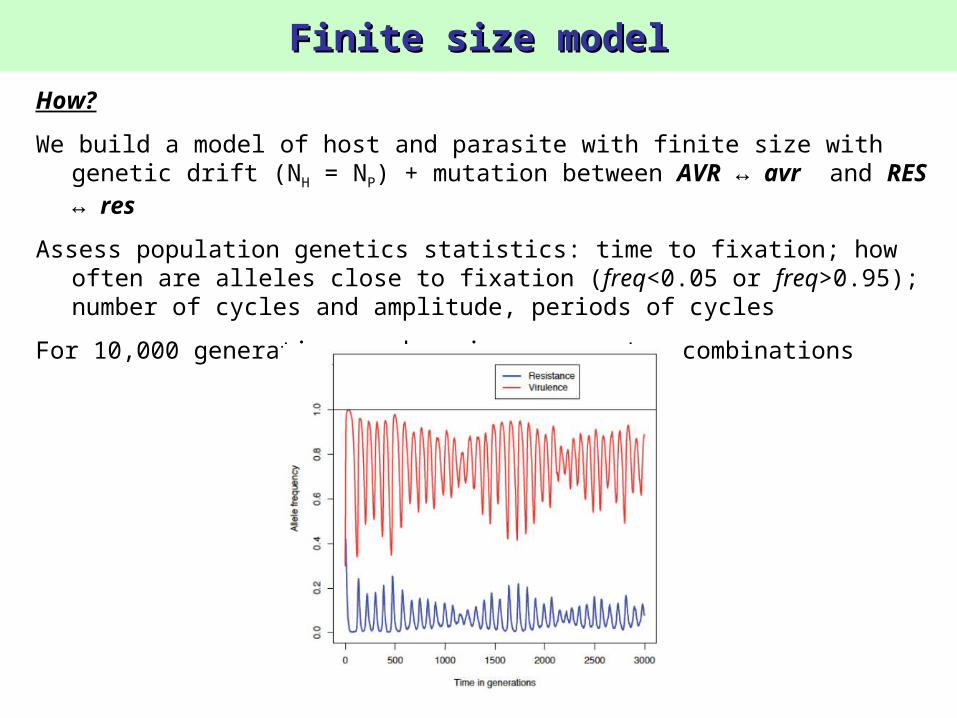
How?
We build a model of host and parasite with finite size with genetic drift (NH = NP) + mutation between AVR ↔ avr and RES ↔ res
Assess population genetics statistics: time to fixation; how often are alleles close to fixation (freq<0.05 or freq>0.95); number of cycles and amplitude, periods of cycles
For 10,000 generations and various parameter combinations
Finite size modelFinite size model
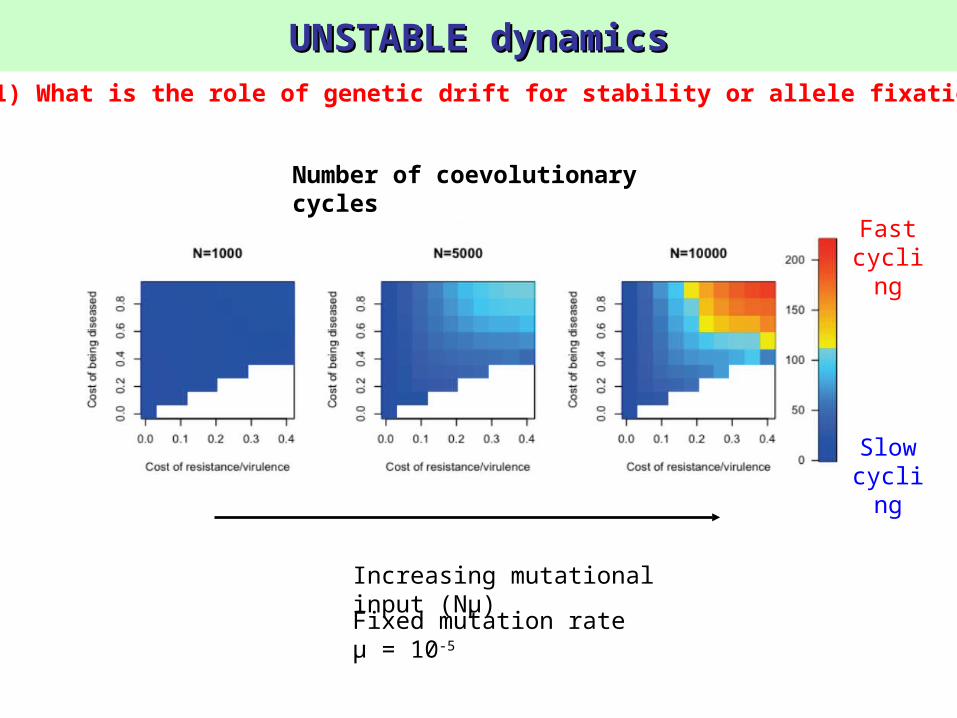
UNSTABLE dynamicsUNSTABLE dynamics
Fixed mutation rate μ = 10-5
Increasing mutational input (Nμ)
1) What is the role of genetic drift for stability or allele fixation?
Number of coevolutionary cycles
Fast cyclin
g
Slow cyclin
g
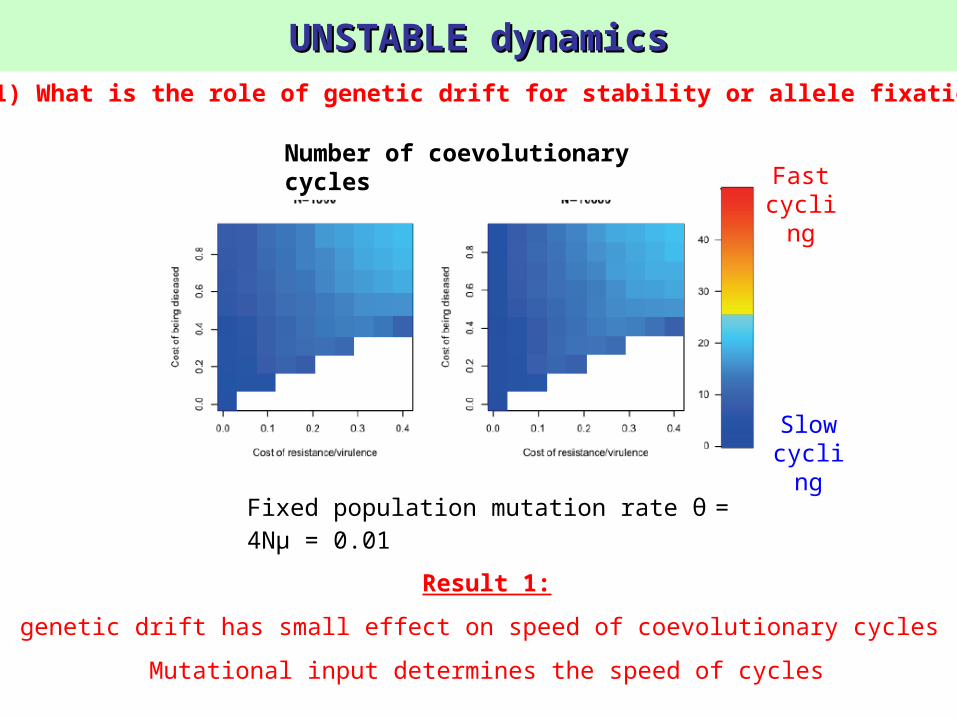
UNSTABLE dynamicsUNSTABLE dynamics
Fixed population mutation rate θ = 4Nμ = 0.01
1) What is the role of genetic drift for stability or allele fixation?
Result 1:
genetic drift has small effect on speed of coevolutionary cycles
Mutational input determines the speed of cycles
Number of coevolutionary cycles Fast
cycling
Slow cyclin
g
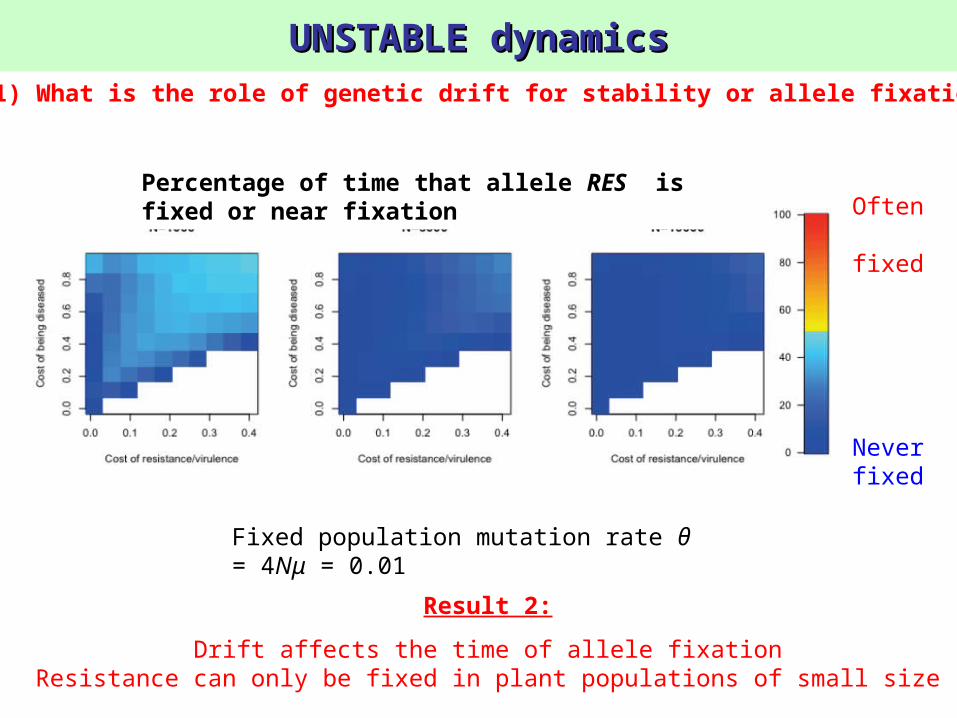
UNSTABLE dynamicsUNSTABLE dynamics
Fixed population mutation rate θ = 4Nμ = 0.01
1) What is the role of genetic drift for stability or allele fixation?
Result 2:
Drift affects the time of allele fixationResistance can only be fixed in plant populations of small size
Percentage of time that allele RES is fixed or near fixation Often
fixed
Never fixed
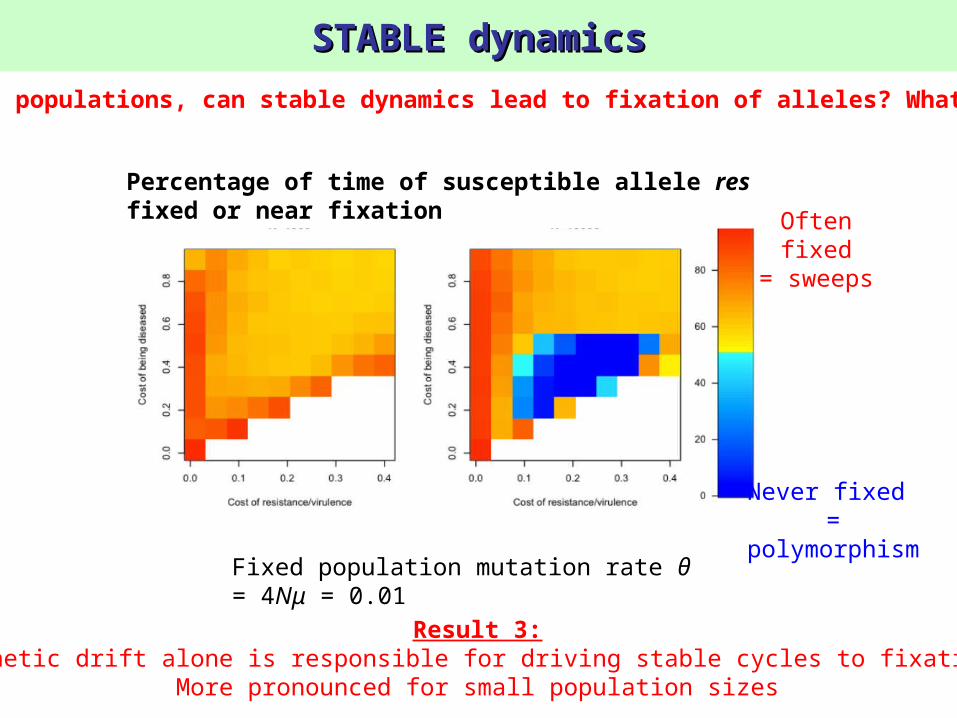
STABLE dynamicsSTABLE dynamics
Fixed population mutation rate θ = 4Nμ = 0.01
Result 3:Genetic drift alone is responsible for driving stable cycles to fixation
More pronounced for small population sizes
Percentage of time of susceptible allele res fixed or near fixation
Never fixed =
polymorphism
Often fixed
= sweeps
2) In finite populations, can stable dynamics lead to fixation of alleles? What conditions?
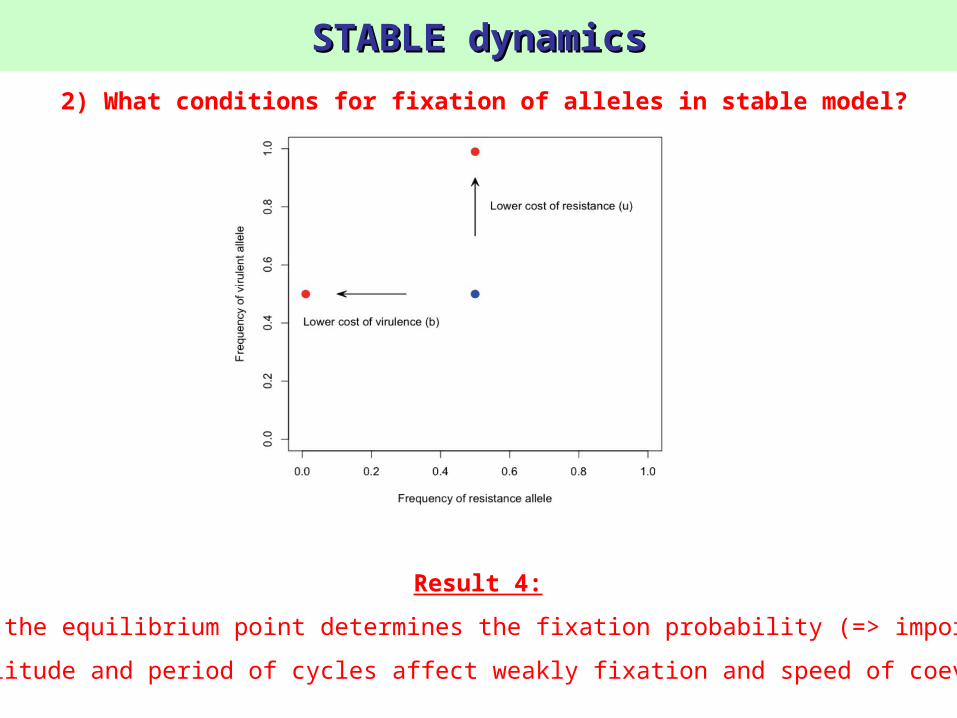
STABLE dynamicsSTABLE dynamics
Result 4:
The location of the equilibrium point determines the fixation probability (=> importance of costs)
BUT amplitude and period of cycles affect weakly fixation and speed of coevolution
2) What conditions for fixation of alleles in stable model?
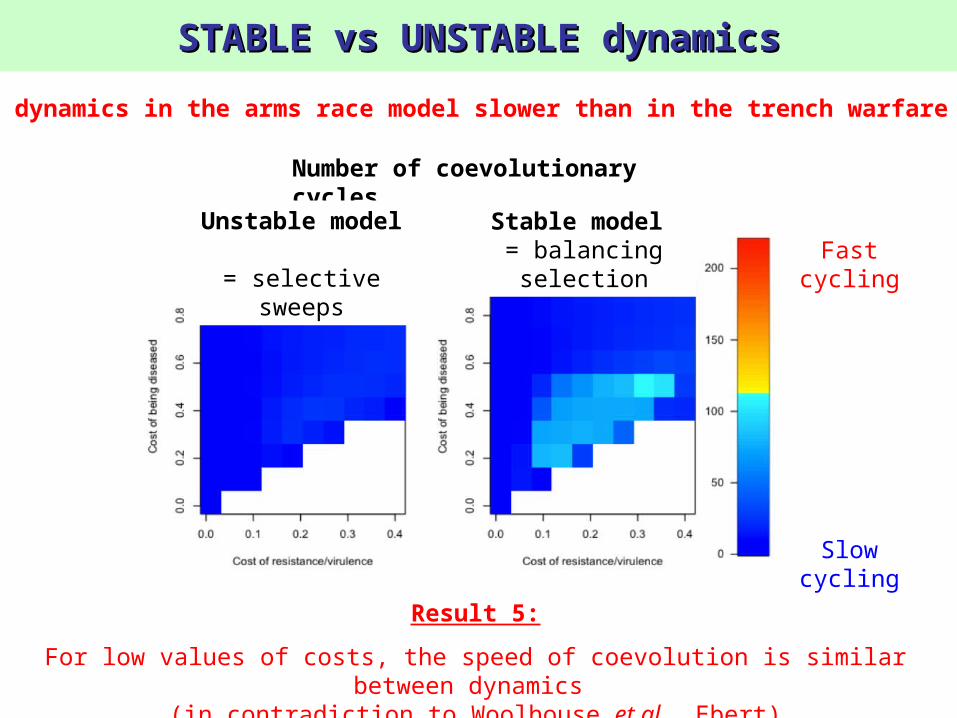
STABLE vs UNSTABLE dynamicsSTABLE vs UNSTABLE dynamics
Result 5:
For low values of costs, the speed of coevolution is similar between dynamics
(in contradiction to Woolhouse et al., Ebert)
3) Are dynamics in the arms race model slower than in the trench warfare model?
Number of coevolutionary cycles
Fast cycling
Slow cycling
Unstable model
= selective sweeps
Stable model = balancing
selection
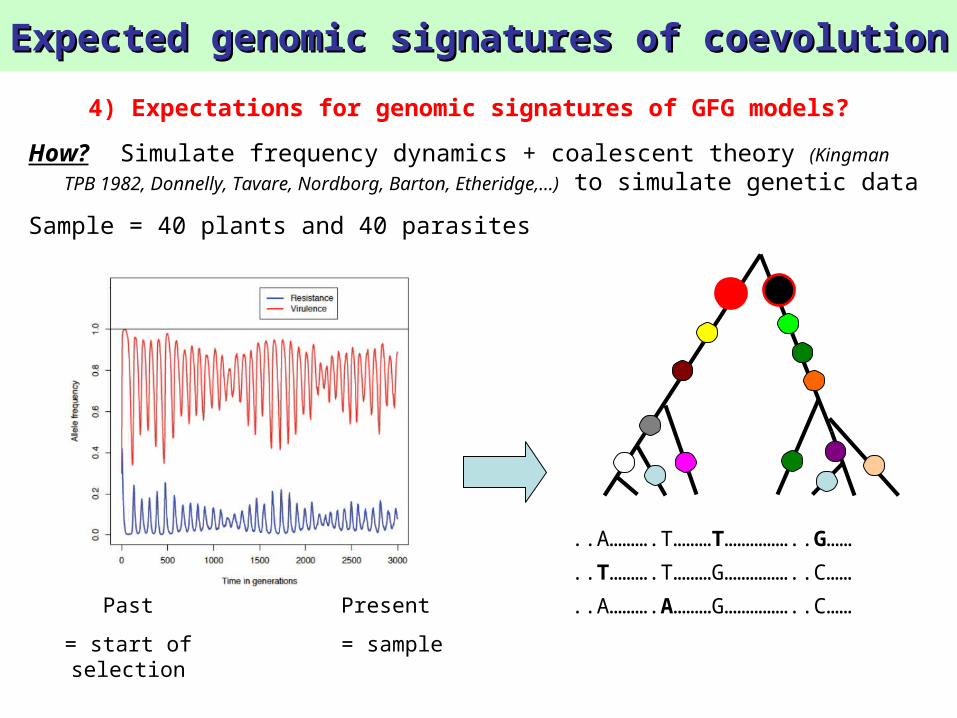
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
4) Expectations for genomic signatures of GFG models?
How? Simulate frequency dynamics + coalescent theory (Kingman TPB
1982, Donnelly, Tavare, Nordborg, Barton, Etheridge,…) to simulate genetic data
Sample = 40 plants and 40 parasites
..A……….T………T……………..G……
..T……….T………G……………..C……
..A……….A………G……………..C……
Present
= sample
Past
= start of selection
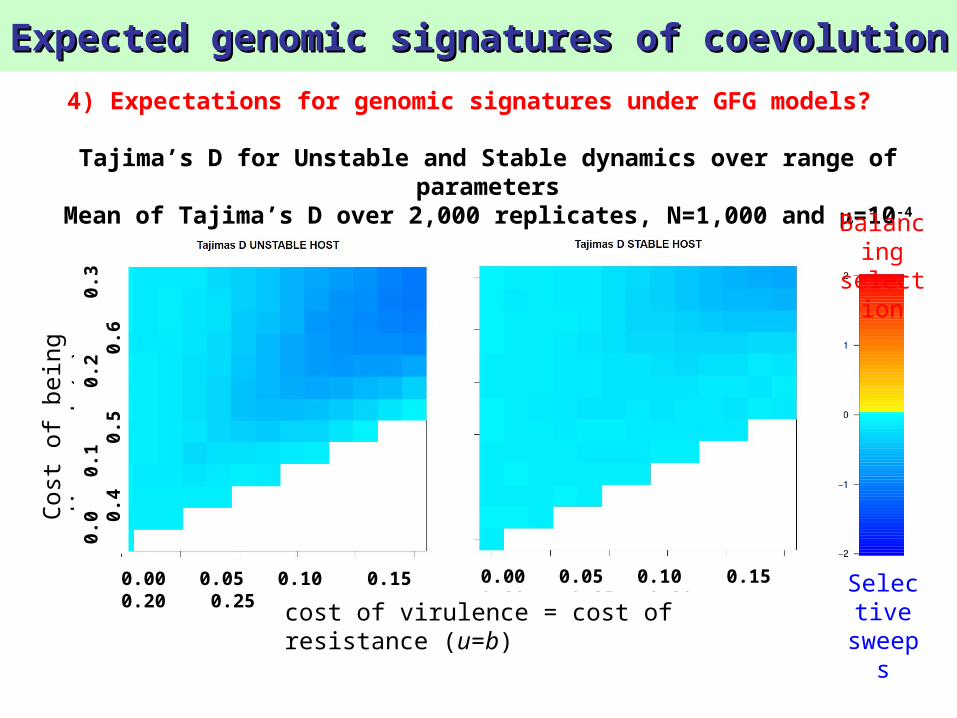
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
Tajima’s D for Unstable and Stable dynamics over range of parameters
Mean of Tajima’s D over 2,000 replicates, N=1,000 and =10-4
4) Expectations for genomic signatures under GFG models?
Balancing
selection
Selective
sweeps
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30
cost of virulence = cost of resistance (u=b)
Cost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
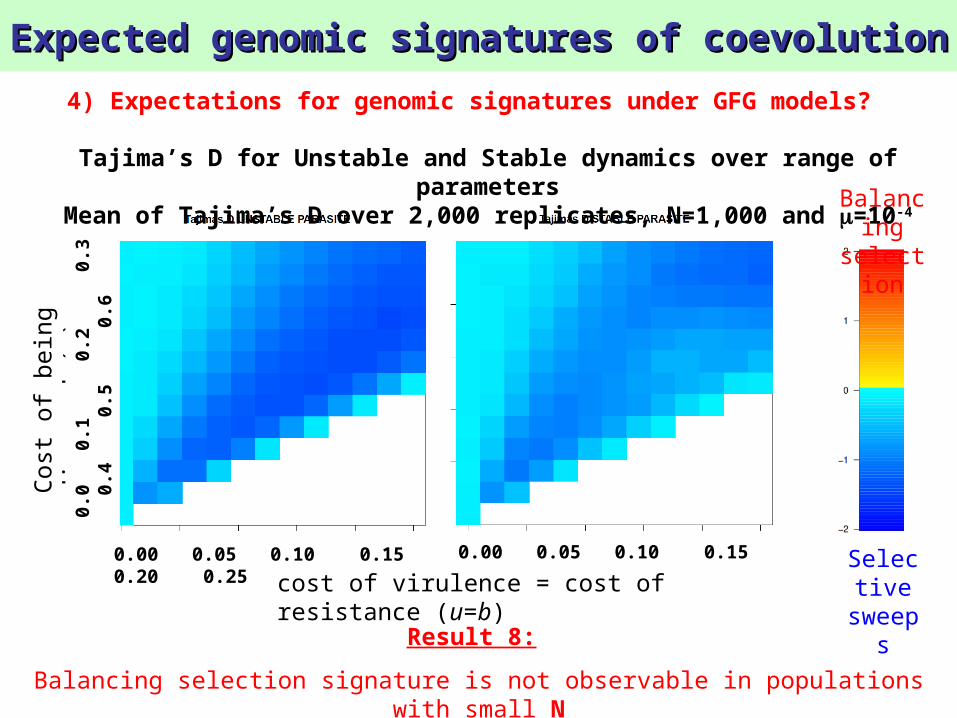
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
4) Expectations for genomic signatures under GFG models?
Cost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
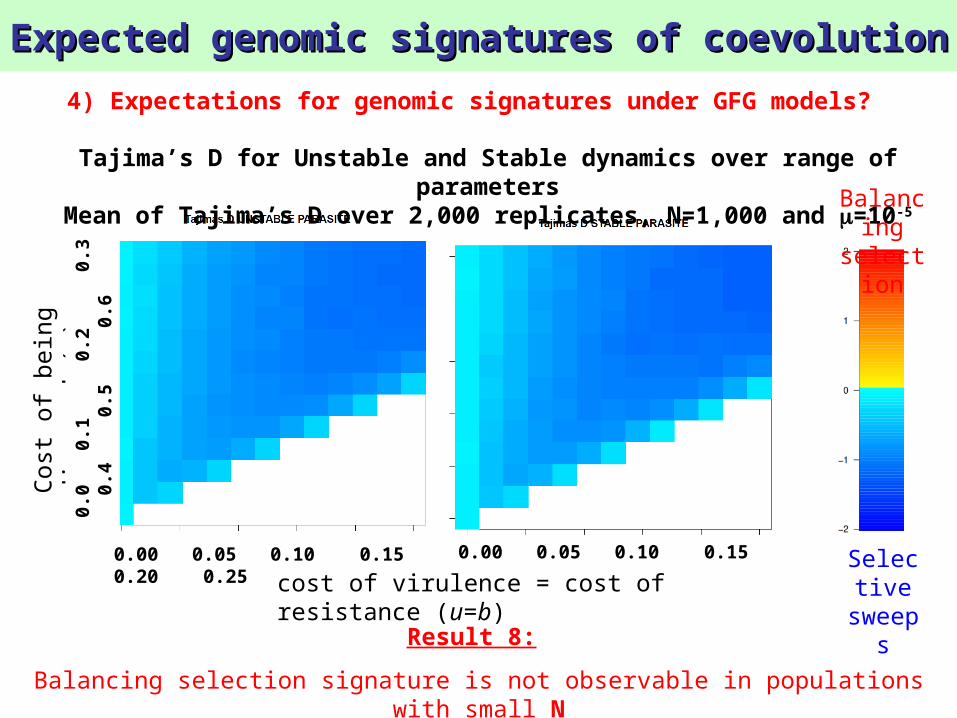
sweepsResult 8:
Balancing selection signature is not observable in populations with small N
Tajima’s D for Unstable and Stable dynamics over range of parameters
Mean of Tajima’s D over 2,000 replicates, N=1,000 and =10-4
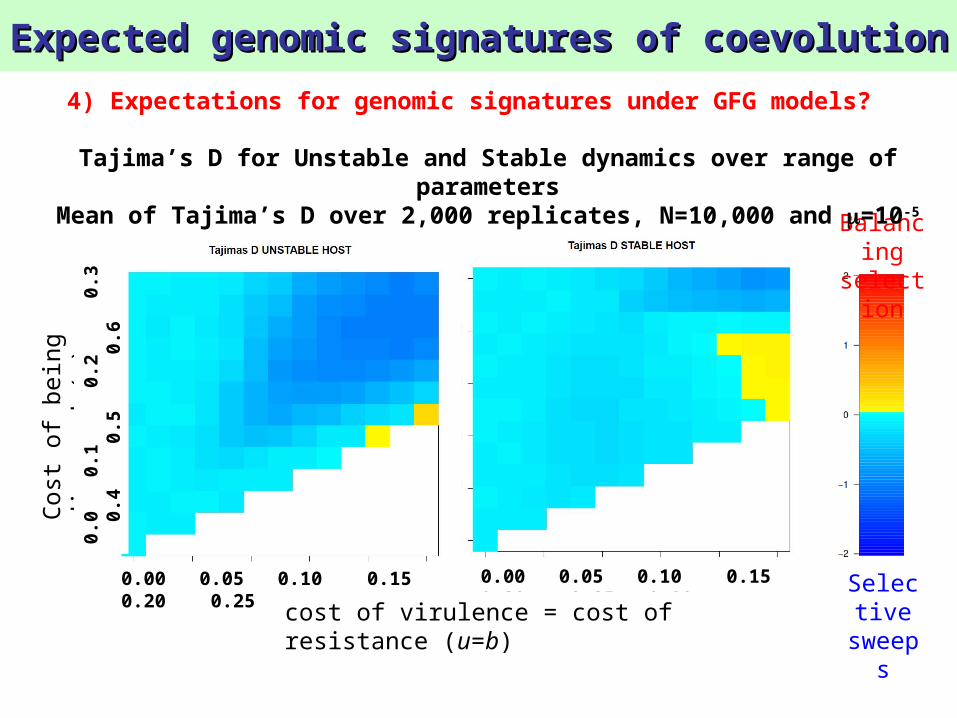
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
4) Expectations for genomic signatures under GFG models?
Balancing
selection
Selective
sweeps
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30
cost of virulence = cost of resistance (u=b)
Cost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
Tajima’s D for Unstable and Stable dynamics over range of parameters
Mean of Tajima’s D over 2,000 replicates, N=10,000 and =10-5
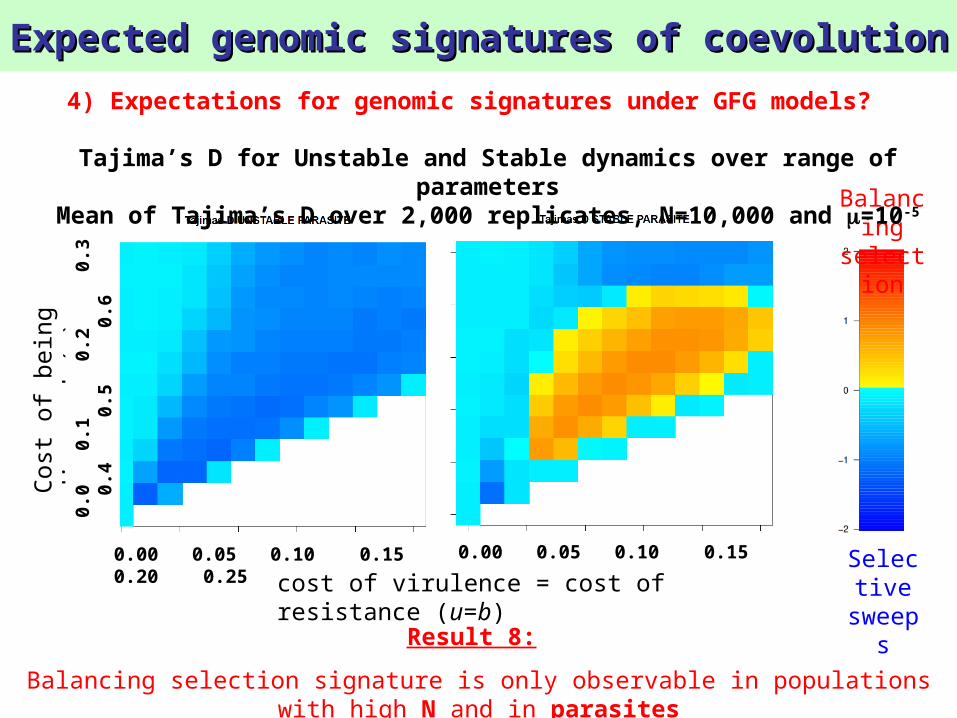
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
4) Expectations for genomic signatures under GFG models?
Cost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweepsResult 8:
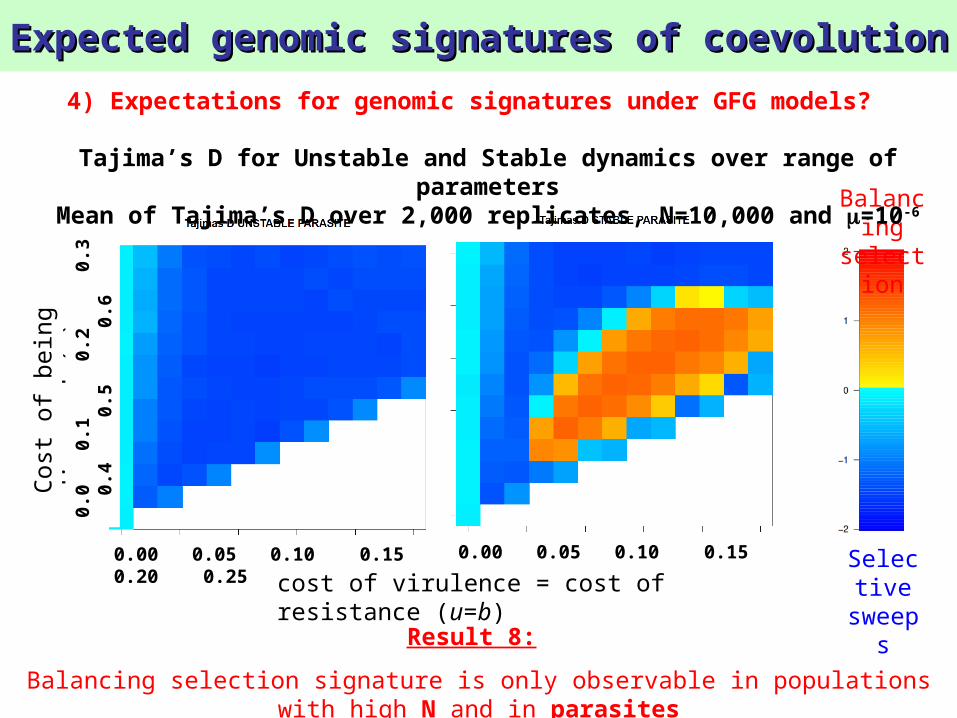
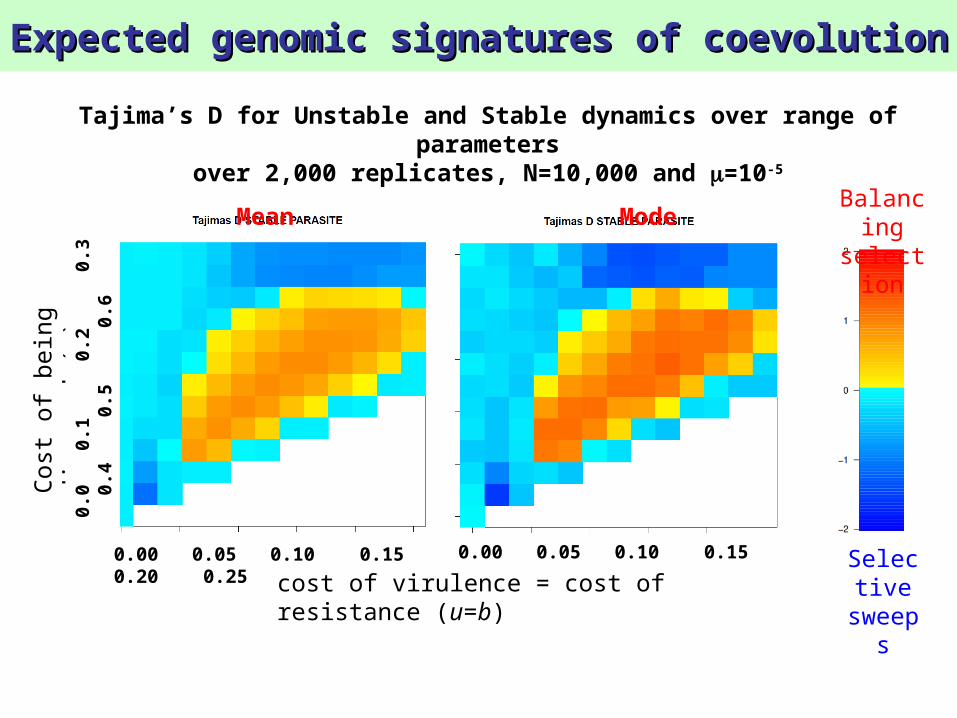
Balancing selection signature is only observable in populations with high N and in parasites
Tajima’s D for Unstable and Stable dynamics over range of parameters
Mean of Tajima’s D over 2,000 replicates, N=10,000 and =10-5
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
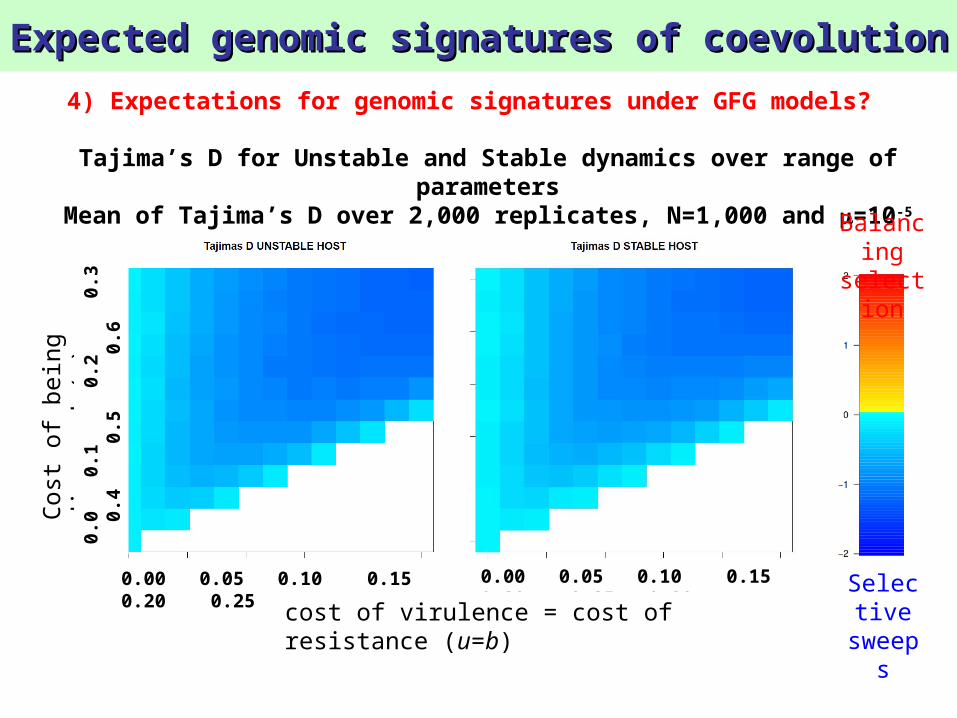
Tajima’s D for Unstable and Stable dynamics over range of parameters
Mean of Tajima’s D over 2,000 replicates, N=1,000 and =10-5
4) Expectations for genomic signatures under GFG models?
Balancing
selection
Selective
sweeps
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30
cost of virulence = cost of resistance (u=b)
Cost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
4) Expectations for genomic signatures under GFG models?
Cost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweepsResult 8:
Balancing selection signature is not observable in populations with small N
Tajima’s D for Unstable and Stable dynamics over range of parameters
Mean of Tajima’s D over 2,000 replicates, N=1,000 and =10-5
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
4) Expectations for genomic signatures under GFG models?
Balancing
selection
Selective
sweeps
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30
cost of virulence = cost of resistance (u=b)
Cost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
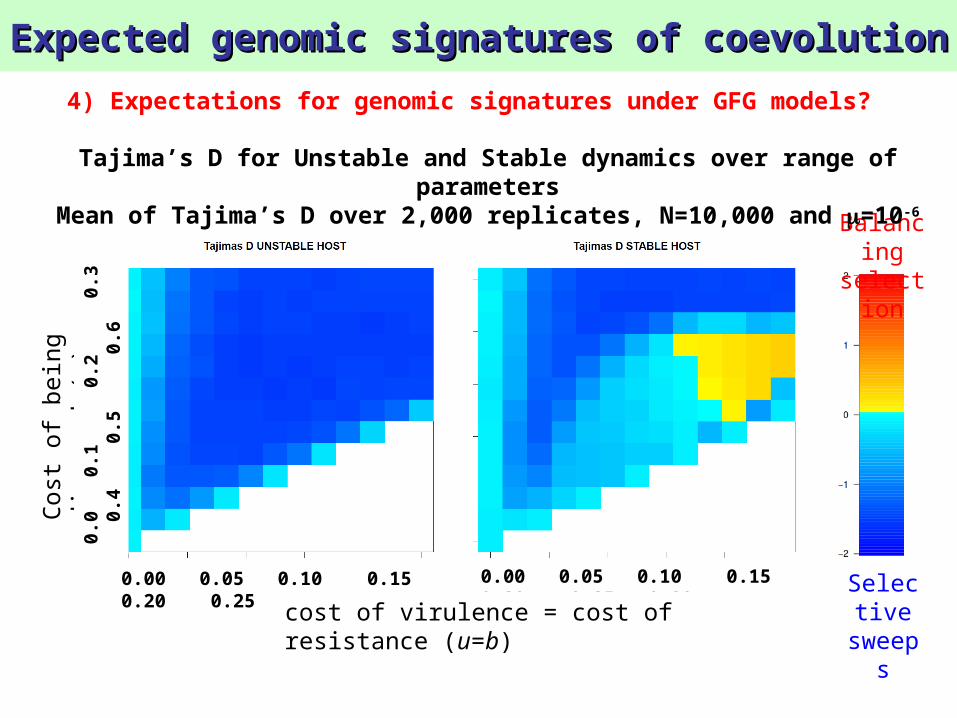
Tajima’s D for Unstable and Stable dynamics over range of parameters
Mean of Tajima’s D over 2,000 replicates, N=10,000 and =10-6
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
4) Expectations for genomic signatures under GFG models?
Cost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweepsResult 8:
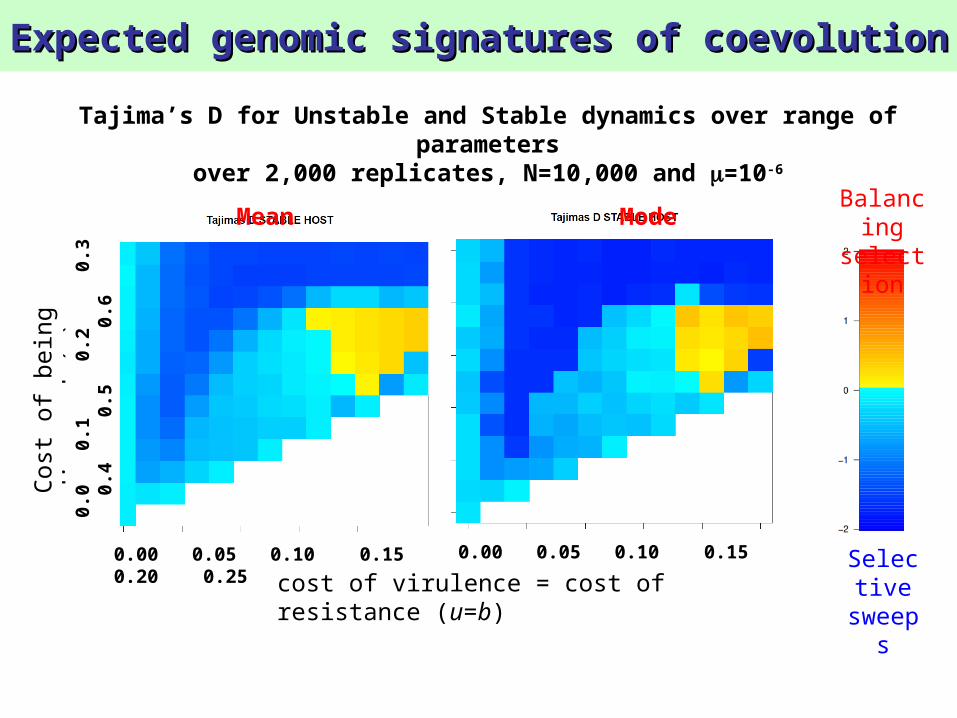
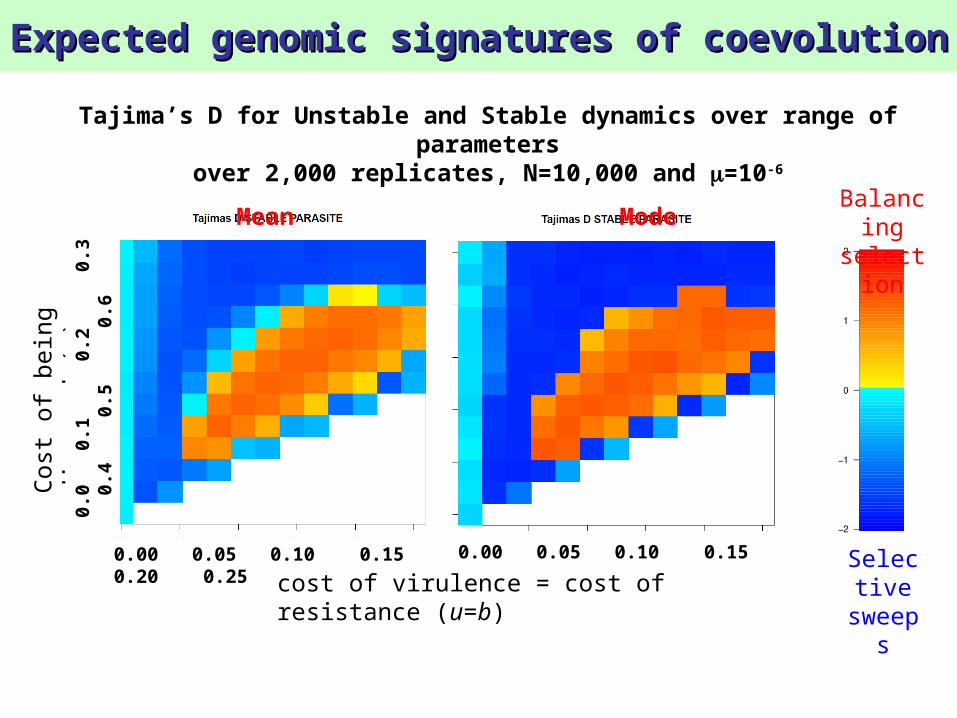
Balancing selection signature is only observable in populations with high N and in parasites
Tajima’s D for Unstable and Stable dynamics over range of parameters
Mean of Tajima’s D over 2,000 replicates, N=10,000 and =10-6
Expected genomic signatures of coevolutionExpected genomic signatures of coevolutionC
ost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweeps
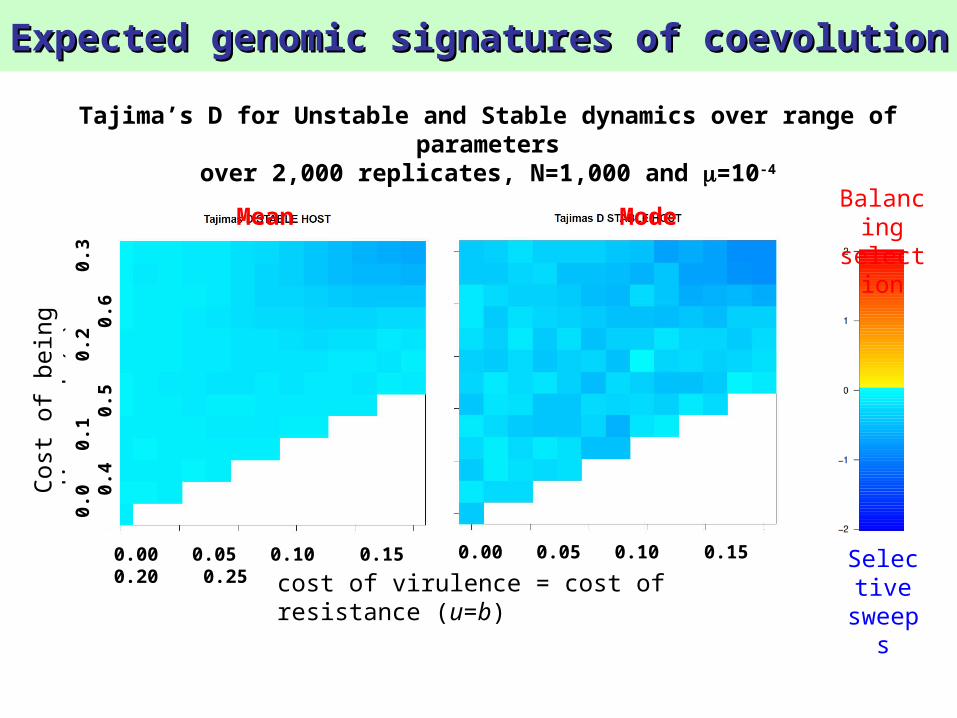
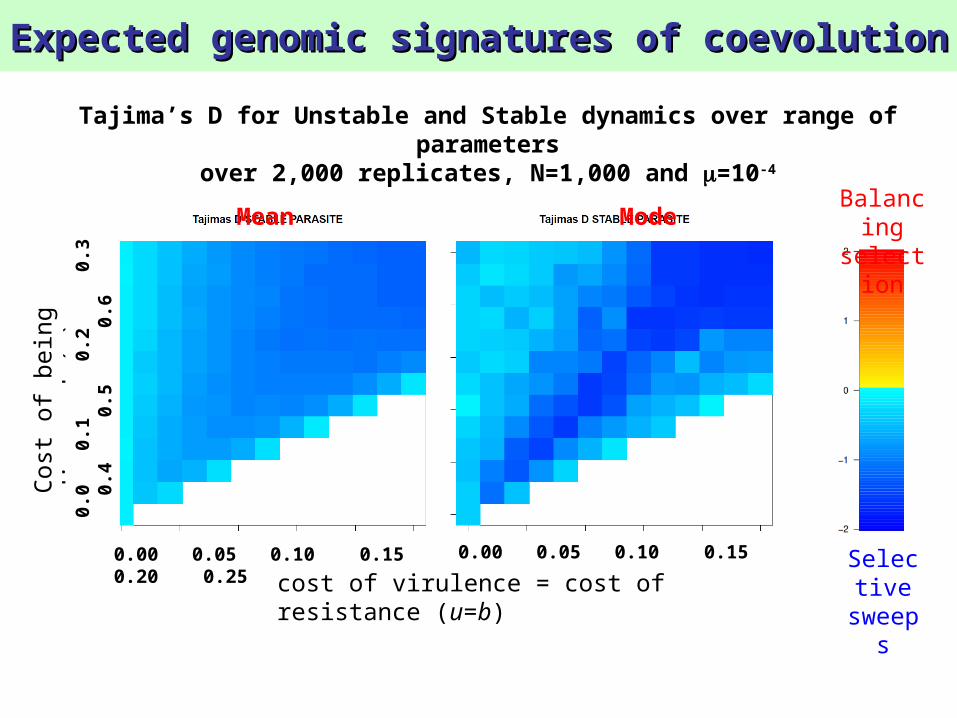
Tajima’s D for Unstable and Stable dynamics over range of parameters
over 2,000 replicates, N=1,000 and =10-4
Mean Mode
Expected genomic signatures of coevolutionExpected genomic signatures of coevolutionC
ost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweeps
Tajima’s D for Unstable and Stable dynamics over range of parameters
over 2,000 replicates, N=1,000 and =10-4
Mean Mode
Expected genomic signatures of coevolutionExpected genomic signatures of coevolutionC
ost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweeps
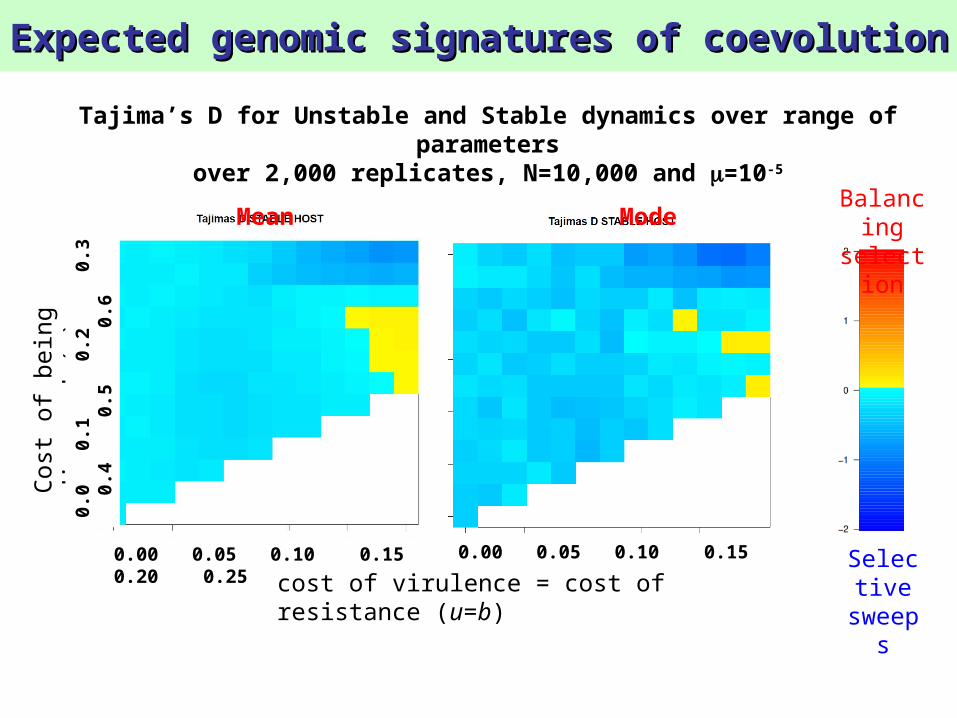
Tajima’s D for Unstable and Stable dynamics over range of parameters
over 2,000 replicates, N=10,000 and =10-5
Mean Mode
Expected genomic signatures of coevolutionExpected genomic signatures of coevolutionC
ost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweeps
Tajima’s D for Unstable and Stable dynamics over range of parameters
over 2,000 replicates, N=10,000 and =10-5
Mean Mode
Expected genomic signatures of coevolutionExpected genomic signatures of coevolutionC
ost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweeps
Tajima’s D for Unstable and Stable dynamics over range of parameters
over 2,000 replicates, N=10,000 and =10-6
Mean Mode
Expected genomic signatures of coevolutionExpected genomic signatures of coevolutionC
ost
of
bein
g
dis
ease
d (
s)0.0
0
.1
0
.2
0
.3
0.4
0.5
0.6
0.00 0.05 0.10 0.15 0.20 0.25 0.30
0.00 0.05 0.10 0.15 0.20 0.25 0.30cost of virulence = cost of
resistance (u=b)
Balancing
selection
Selective
sweeps
Tajima’s D for Unstable and Stable dynamics over range of parameters
over 2,000 replicates, N=10,000 and =10-6
Mean Mode

ConclusionsConclusions
1) What is the role of genetic drift for stability or allele fixation?
2) In finite populations, can stable dynamics lead to fixation of alleles? Under which conditions?
Genetic drift does not influence the speed of coevolution, but the fixation rate of alleles
Alleles can get fixed in a stable model (contrary to common wisdom)
Resistance alleles only get fixed in small populations!!! Diversifying selection at Rgenes must have occurred in small plant populations3) Are dynamics in the arms race model slower than in the trench warfare model?
No, for most parameter values there is no difference between the speed of coevolution
Relevance for field studies following changes in allele frequencies 4) Expectations for genomic signatures under GFG models?
Unstable dynamics can lead to a big variation in genomic signatures
Bad news: almost no chance to find balancing selection in genome scans in plants (ex. Arabidopsis thaliana), AND costs determine the equilibrium point
Good news: genome scans can be used to find balancing selection in parasites

Thanks to hazards of migrationThanks to hazards of migration
Wolfgang Stephan Stefan Laurent Daniel Živković
Mamadou Mboup Katharina Böndel
James Brown
T. Giraud (University Paris 13)
J. Enjalbert (INRA, France)A.L. Laine (University
Helsinki)
Anja HörgerThomas Städler
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
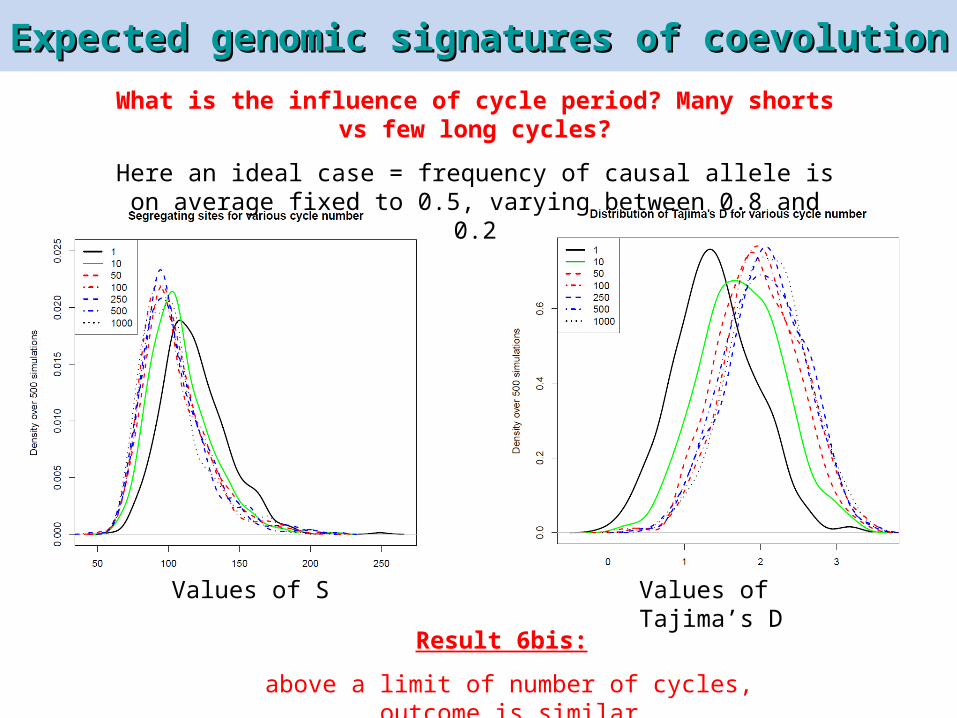
What is the influence of cycle period? Many shorts vs few long cycles?
Here an ideal case = frequency of causal allele is on average fixed to 0.5, varying between 0.8 and 0.2
Values of S Values of Tajima’s D
Result 6bis:
above a limit of number of cycles, outcome is similar
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
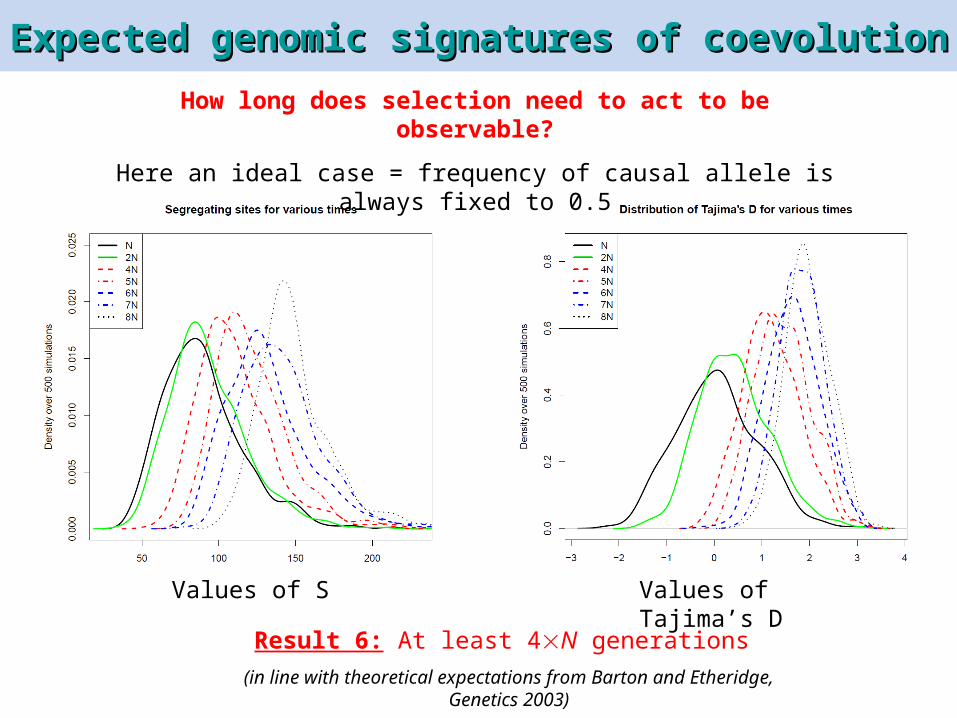
How long does selection need to act to be observable?
Here an ideal case = frequency of causal allele is always fixed to 0.5
Values of S Values of Tajima’s D
Result 6: At least 4N generations
(in line with theoretical expectations from Barton and Etheridge, Genetics 2003)
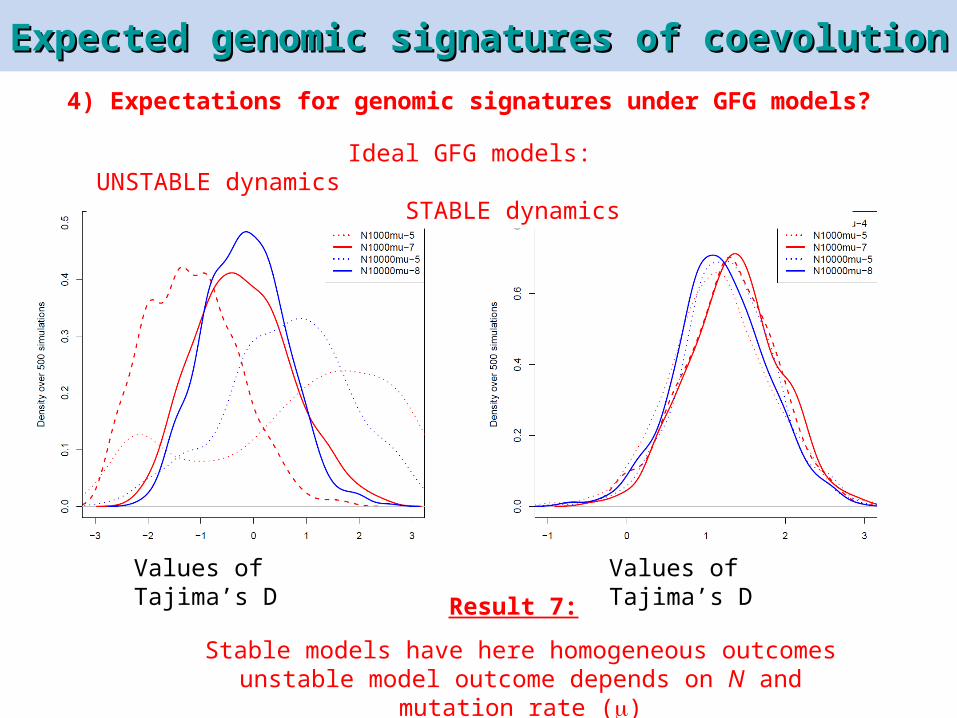
Expected genomic signatures of coevolutionExpected genomic signatures of coevolution
Ideal GFG models:UNSTABLE dynamics STABLE
dynamics
Values of Tajima’s DResult 7:
Stable models have here homogeneous outcomesunstable model outcome depends on N and mutation
rate ()
Values of Tajima’s D
4) Expectations for genomic signatures under GFG models?
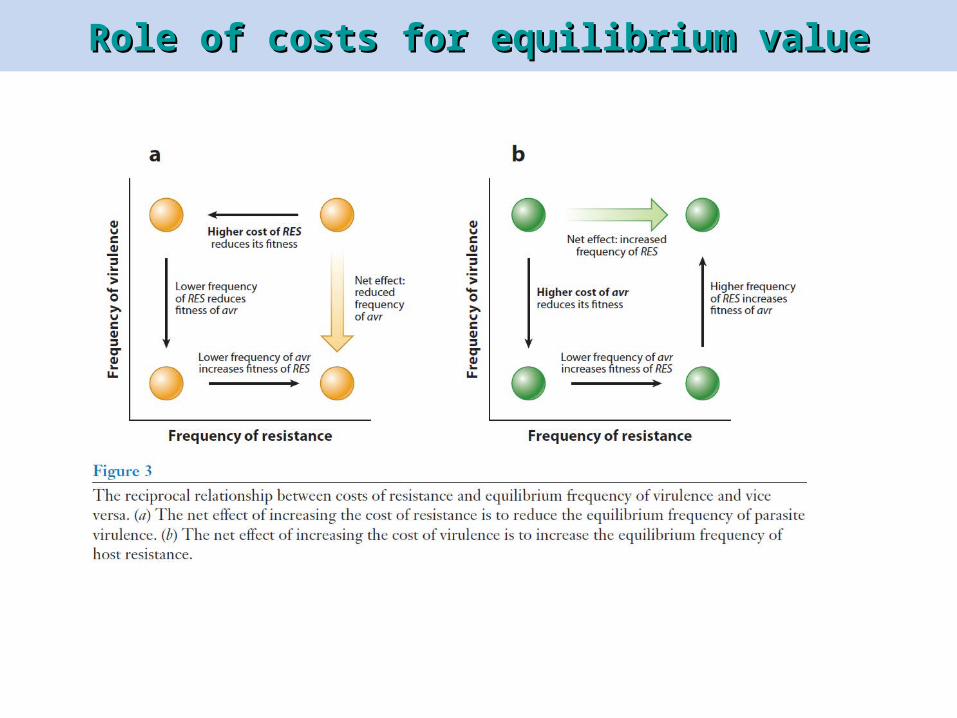
Role of costs for equilibrium valueRole of costs for equilibrium value
Related Documents
![The wild tomato species [i]Solanum chilense[i] …39 Background 40 41 In nature, plants are exposed to a wide range of pathogens and pests. While in most cases the 42 plants appear](https://static.cupdf.com/doc/110x72/5e9f57e45db9ad41fc5db019/the-wild-tomato-species-isolanum-chilensei-39-background-40-41-in-nature-plants.jpg)










