Proc. Nat. Acad. Sci. USA Vol. 70, No. 11, pp. 323.5-3239, November 1973 Membrane Optical Activity: Some Facts and Fallacies (light scattering/poly(L-lysine)/circular dichroism) DONALD F. H. WALLACH*t, DAVID A. LOW*, AND ALEXANDER V. BERTLANDt * Tufts-New England Medical Center, Boston, Massachusetts 02111 USA; and t Merck Institute for Therapeutic Research, West Point, Pennsylvania USA Communicated by J. L. Oncley, May 30, 1973 ABSTRACT The circular dichroism of hypothetical, water-filled, spherical shells, 75-3500 nm in radius, with walls 7.5 nm thick, composed of poly(L-lysine) in various conformational proportions, and suspended in water, were computed from the known optical properties of this polypeptide by classical general light-scattering theory (Mie theory). Comparison of the computed curves of circular dichroism spectra with those of diverse mem- branes reveals large discrepancies below 215 nm and shows that light scattering does not adequately account for the optical activity of membranes containing appreciable proportions of nonhelical conformation. However, tur- bidity effects can explain the anomalies of membrane optical rotatory dispersion near 233 nm, if not uniquely so. We conclude that the optical activity of neither most soluble proteins nor membrane proteins can provide accurate conformational information when synthetic polypeptides are used as standards and list the reasons for this argument. We also show that present techniques to "correct" membrane optical activity are likely to produce additional artifact. One approach to the study of membrane-protein architecture involves the use of spectroscopic methods. However, no single technique can derive unambiguous data from large molecules or molecular aggregates. This is particularly true for optical activity measurements, which have found wide application in attempts to extract conformational information from soluble and membrane proteins, with synthetic polypeptides as reference standards. Indeed comparison of the circular di- chroism (CD) and optical rotatory dispersion (ORD) of most proteins with those characterizing synthetic homopolypeptides with a-helical, (3-, and unordered structures, is unlikely to yield satisfactory conformation analyses of the proteins. The reasons are as follows: (a) 'Many proteins (1-4), including membrane components (5-8), contain significant and sometimes functionally variable proportions of (-conformation; one must therefore analyze unknown substances in terms of three conformations. This is indeed difficult, even with a single synthetic polypeptide as standard, since seven optically active chromophores are concerned: these are the n-7r* transitions of the three confor- mations, the 7r-7r* transitions of the "unordered" and (3- structures, the I -polarized helical 7r- r* transition and the Il-polarized 7r- r* transition. Additional complications arise from band overlap and uncertainty whether the bands have the frequencies, intensities, widths, and shapes found in the usual polypeptide models. (b) a-Helical optical activity increases with the number of consecutive helically arrayed residues up to about 20 (9-11) and decreases with deviation from ideal a-helicity; but x-ray analyses of many globular proteins show that there are usually fewer than 20 residues per helical segment (1-3, 12) and that distortions are common. (c) Even in synthetic polypeptides the optical activity of a-helices is slightly (13) and that of the (3-conformation (14, 15) strongly side-chain dependent. (d) "Unordered" segments in proteins, if irregular, are relatively fixed and structured and vary from protein to protein. They are not statistically distributed elements of varying structure or size. They can also not be assumed to resemble the "random" states of poly(-glutamate) or poly(L- lysine) which have elements of local extended helix (16-19). Further, the "infinite" pleated sheet structure formed by some polypeptides appears inadequate as a model for less extended d-structures found in proteins. (e) X-ray crystallographic (12) and probe experiments (20) suggest that many of the peptide linkages of globular proteins lie in an apolar, highly polarizable environment, unlike those of reference polypeptides in water solution. This environment mav reduce rotational strengths and shift the frequencies of the optically active bands (21). (f) Conformations other than the conventional right- handed a-helical, f3-, and "unoidered" structures must be considered. Thus, although the left-handed a-helix is intrinsi- cally less stable than the right-handed helix, other factors may sometimes favor it (22, 23). Thus polyv(-benzyl--aspartate) forms a right-handed a-helix wheii spread at air: water inter- faces, but exists either as unordered structures or left-handed helices in solution (23). These findings should be kept in mind, since membranes may contain polar: apolar boundaries such as exemplified by an air: water interface. (g) One cannot properly assess the contribution of side- chain optical activity of globular proteins to their CD and ORD in the "peptide region" (19). Awareness of these fundamental problems has fostered the search for new means, including the use of protein standards, to extract unambiguous conformational information from optical activity measurements (19, 21, 24-26). The above complications indubitably apply also to mem- brane proteins (27), but have been largely ignored in the extensive reports on light-scattering artifacts which fail to rigorously analyze membrane optical activity over the full range of peptide absorption accessible. We will discuss this matter below, but stress first that (i) different membranes can vary greatly in their morphological, chemical, and physical proper- Abbreviations: CD, circular dichroisin; ORD, optical rotatory dispersion. I To whom correspondence should be addressed. 3235

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Proc. Nat. Acad. Sci. USAVol. 70, No. 11, pp. 323.5-3239, November 1973
Membrane Optical Activity: Some Facts and Fallacies(light scattering/poly(L-lysine)/circular dichroism)
DONALD F. H. WALLACH*t, DAVID A. LOW*, AND ALEXANDER V. BERTLANDt
* Tufts-New England Medical Center, Boston, Massachusetts 02111 USA; and t Merck Institute for Therapeutic Research, West Point,Pennsylvania USA
Communicated by J. L. Oncley, May 30, 1973
ABSTRACT The circular dichroism of hypothetical,water-filled, spherical shells, 75-3500 nm in radius, withwalls 7.5 nm thick, composed of poly(L-lysine) in variousconformational proportions, and suspended in water,were computed from the known optical properties of thispolypeptide by classical general light-scattering theory(Mie theory). Comparison of the computed curves ofcircular dichroism spectra with those of diverse mem-branes reveals large discrepancies below 215 nm and showsthat light scattering does not adequately account for theoptical activity of membranes containing appreciableproportions of nonhelical conformation. However, tur-bidity effects can explain the anomalies of membraneoptical rotatory dispersion near 233 nm, if not uniquely so.We conclude that the optical activity of neither mostsoluble proteins nor membrane proteins can provideaccurate conformational information when syntheticpolypeptides are used as standards and list the reasons forthis argument. We also show that present techniques to"correct" membrane optical activity are likely to produceadditional artifact.
One approach to the study of membrane-protein architectureinvolves the use of spectroscopic methods. However, no singletechnique can derive unambiguous data from large moleculesor molecular aggregates. This is particularly true for opticalactivity measurements, which have found wide application inattempts to extract conformational information from solubleand membrane proteins, with synthetic polypeptides asreference standards. Indeed comparison of the circular di-chroism (CD) and optical rotatory dispersion (ORD) of mostproteins with those characterizing synthetic homopolypeptideswith a-helical, (3-, and unordered structures, is unlikely toyield satisfactory conformation analyses of the proteins. Thereasons are as follows:
(a) 'Many proteins (1-4), including membrane components(5-8), contain significant and sometimes functionally variableproportions of (-conformation; one must therefore analyzeunknown substances in terms of three conformations. This isindeed difficult, even with a single synthetic polypeptide asstandard, since seven optically active chromophores areconcerned: these are the n-7r* transitions of the three confor-mations, the 7r-7r* transitions of the "unordered" and (3-structures, the I -polarized helical 7r- r* transition and theIl-polarized 7r- r* transition. Additional complications arisefrom band overlap and uncertainty whether the bands havethe frequencies, intensities, widths, and shapes found in theusual polypeptide models.
(b) a-Helical optical activity increases with the number ofconsecutive helically arrayed residues up to about 20 (9-11)and decreases with deviation from ideal a-helicity; but x-rayanalyses of many globular proteins show that there areusually fewer than 20 residues per helical segment (1-3, 12)and that distortions are common.
(c) Even in synthetic polypeptides the optical activity ofa-helices is slightly (13) and that of the (3-conformation (14,15) strongly side-chain dependent.
(d) "Unordered" segments in proteins, if irregular, arerelatively fixed and structured and vary from protein toprotein. They are not statistically distributed elements ofvarying structure or size. They can also not be assumed toresemble the "random" states of poly(-glutamate) or poly(L-lysine) which have elements of local extended helix (16-19).Further, the "infinite" pleated sheet structure formed by somepolypeptides appears inadequate as a model for less extendedd-structures found in proteins.
(e) X-ray crystallographic (12) and probe experiments (20)suggest that many of the peptide linkages of globular proteinslie in an apolar, highly polarizable environment, unlike thoseof reference polypeptides in water solution. This environmentmav reduce rotational strengths and shift the frequencies ofthe optically active bands (21).
(f) Conformations other than the conventional right-handed a-helical, f3-, and "unoidered" structures must beconsidered. Thus, although the left-handed a-helix is intrinsi-cally less stable than the right-handed helix, other factors maysometimes favor it (22, 23). Thus polyv(-benzyl--aspartate)forms a right-handed a-helix wheii spread at air: water inter-faces, but exists either as unordered structures or left-handedhelices in solution (23). These findings should be kept in mind,since membranes may contain polar: apolar boundaries suchas exemplified by an air: water interface.
(g) One cannot properly assess the contribution of side-chain optical activity of globular proteins to their CD andORD in the "peptide region" (19).Awareness of these fundamental problems has fostered the
search for new means, including the use of protein standards,to extract unambiguous conformational information fromoptical activity measurements (19, 21, 24-26).The above complications indubitably apply also to mem-
brane proteins (27), but have been largely ignored in theextensive reports on light-scattering artifacts which fail torigorously analyze membrane optical activity over the full rangeof peptide absorption accessible. We will discuss this matterbelow, but stress first that (i) different membranes can varygreatly in their morphological, chemical, and physical proper-
Abbreviations: CD, circular dichroisin; ORD, optical rotatorydispersion.I To whom correspondence should be addressed.
3235
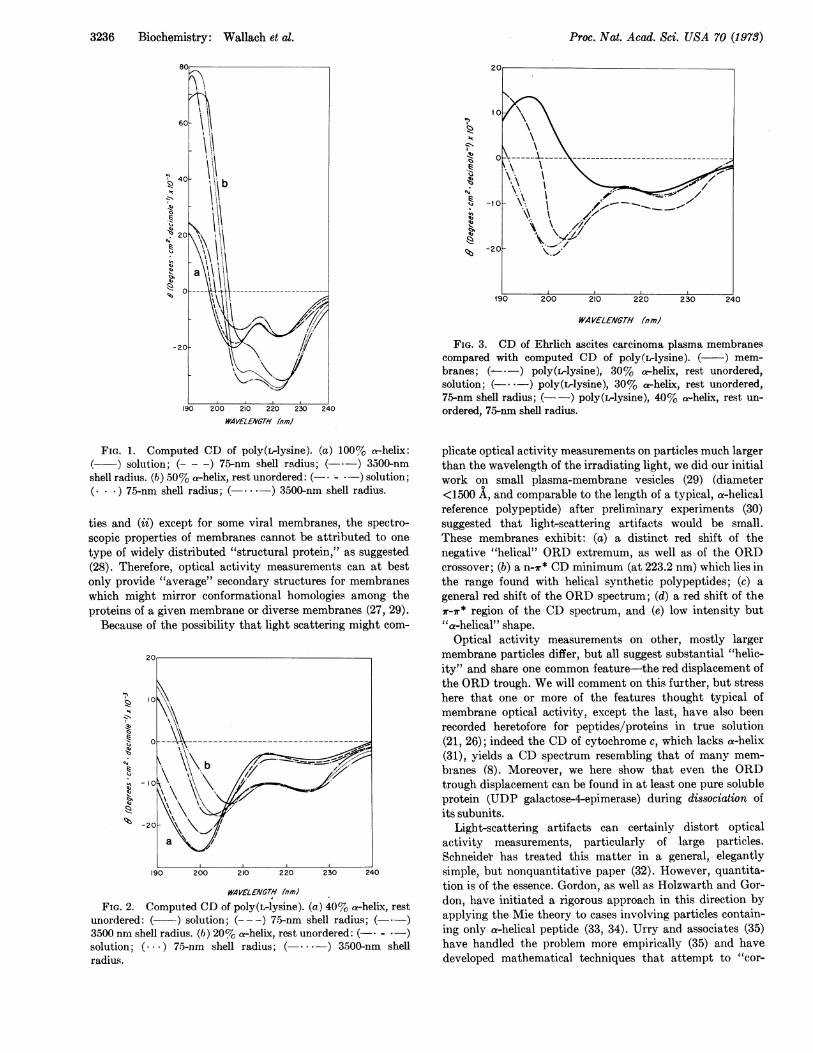
3236 Biochemistry: Wallach et al.
IS%
qua
- 2C
WAVELENGTH (n}m
FIG. 1. Computed CD of poly(ilysine). (a) 100%t a-helix:solution; (---) 75-nm shell radius; (-*-) 3500-nm
shell radius. (b) 50% a-helix, rest unordered: (-* - *-) solution;(. *) 75-nm shell radius; (- -) 3500-nm shell radius.
ties and (ii) except for some viral membranes, the spectro-scopic properties of membranes cannot be attributed to one
type of widely distributed "structural protein," as suggested(28). Therefore, optical activity measurements can at bestonly provide "average" secondary structures for membraneswhich might mirror conformational homologies among theproteins of a given membrane or diverse membranes (27, 29).
Because of the possibility that light scattering might com-
WAVELENGTH (nm)
FIG. 2. Computed CD of poly(L-lysine). (a) 40% a-helix, restunordered: ( -) solution; (--- -) 75-nm shell radius; (- .-)3500 nrn shell radius. (i) 20% a-helix, rest unordered: (-* - *-)solution; ( ... ) 75-nm shell radius; (*. - *-) 3500-nm shellradius.
I-) \\ \0 ----- A.- --\------------------------------- 7:-
-20F
190 200 210 220 230 240
WAVELENGTH (nrml
FIG. 3. CD of Ehrlich ascites carcinoma plasma membranescompared with computed CD of poly(L-lysine). ( ) mem-
branes; (-*-) poly(L-lysine), 30% a-helix, rest unordered,solution; (-. -) poly(L-lysine), 30% a-helix, rest unordered,75-nm shell radius; (- -) poly(ilysine), 40% a-helix, rest un-
ordered, 75-nm shell radius.
plicate optical activity measurements on particles much largerthan the wavelength of the irradiating light, we did our initialwork on small plasma-membrane vesicles (29) (diameter<1500 A, and comparable to the length of a typical, a-helicalreference polypeptide) after preliminary experiments (30)suggested that light-scattering artifacts would be small.These membranes exhibit: (a) a distinct red shift of thenegative "helical" ORD extremum, as well as of the ORDcrossover; (b) a no7r* CD minimum (at 223.2 nm) which lies inthe range found with helical synthetic polypeptides; (c) a
general red shift of the ORD spectrum; (d) a red shift of ther-7r* region of the CD spectrum, and (e) low intensity but'a-helical" shape.Optical activity measurements on other, mostly larger
membrane particles differ, but all suggest substantial "helic-ity" and share one common feature-the red displacement ofthe ORD trough. We will comment on this further, but stresshere that one or more of the features thought typical ofmembrane optical activity, except the last, have also beenrecorded heretofore for peptides/proteins in true solution(21, 26); indeed the CD of cytochrome c, which lacks a-helix(31), yields a CD spectrum resembling that of many mem-
branes (8). Moreover, we here show that even the ORDtrough displacement can be found in at least one pure solubleprotein (UDP galactose-4-epimerase) during dissociation ofits subunits.
Light-scattering artifacts can certainly distort opticalactivity measurements, particularly of large particles.Schneider has treated this matter in a general, elegantlysimple, but nonquantitative paper (32). However, quantita-tion is of the essence. Gordon, as well as Holzwarth and Gor-don, have initiated a rigorous approach in this direction byapplying the Mie theory to cases involving particles contain-ing only a-helical peptide (33, 34). Urry and associates (35)have handled the problem more empirically (35) and havedeveloped mathematical techniques that attempt to "cor-
.-._/~~ ~ ~
.\ I
,\\ ' X
\\
Proc. Nat. Acad. Sci. USA 70 (1978)
-1 o~
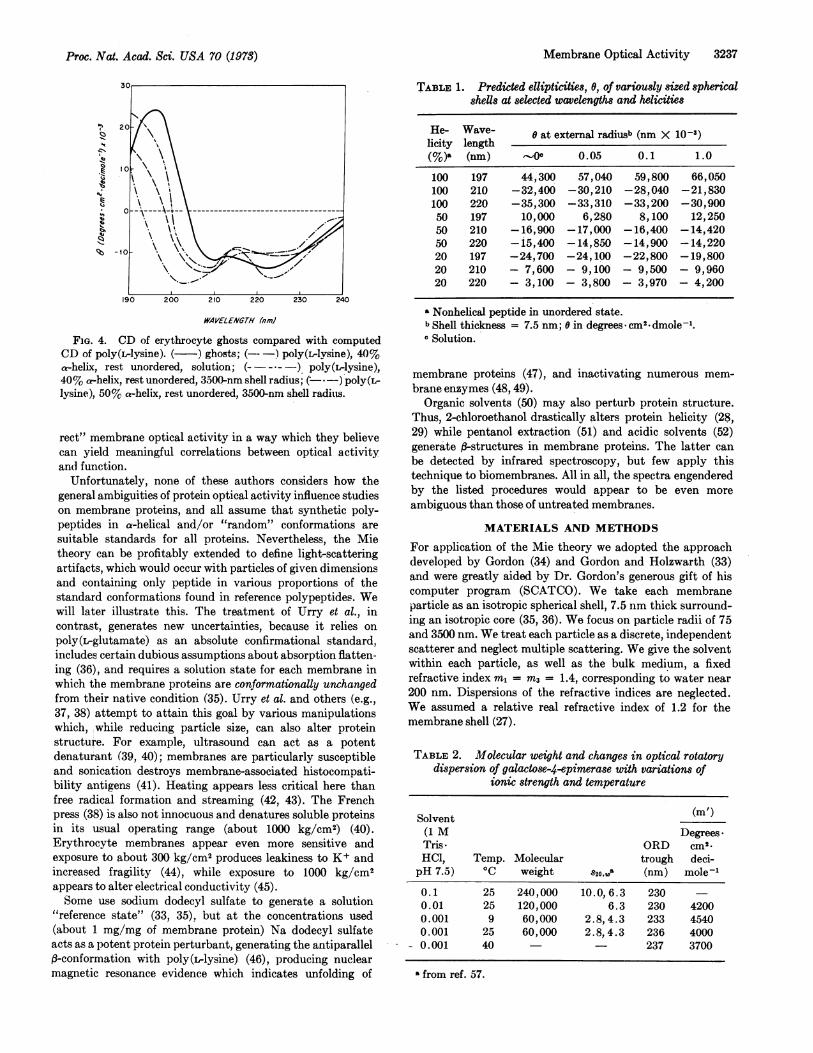
Membrane Optical Activity 3237
20
190 200 210 220 230 240
WAV/ELENGTHf (7mj
FIG. 4. CD of erythrocyte ghosts compared with computedCD of poly(ilysine). ( ) ghosts; ( !poly(LIvlsine), 40)%ahelix, rest unordered, solution; (a )poly(Lrlysine),40 %0 ahelix, refit unordered, 3500-nm shell radius; (I )poly(L-lysine), 50% at-helix, rest unordered, 3500 nm shell radices.
rect" membrane optical activity in a way which they believecan yield meaningful correlations between optical activityandl function.
Unfortunately, none of these authors considers how thegeneral ambiguities of protein optical activity influence studieson membrane proteins, and all assume that synthetic poly-peptides in a-helical and/or "random" conformations aresuitable standards for all proteins. Nevertheless, the Mietheory can be profitably extended to define light-scatteringartifacts, which would occur with particles of given dimensionsand containing only peptide in various proportions of thestandard conformations found in reference polypeptides. Wewill later illustrate this. The treatment of Urry et al., incontrast, generates new uncertainties, because it relies onpoly(L-glutamate) as an absolute confirmational standard,includes certain dubious assumptions about absorption flatten-ing (36) , and requires a solution state for each membrane inwhich the membrane proteins are conformationally unchangedfrom their native condition (35). Urry et al. and others (e.g.,37, 38) attempt to attain this goal by various manipulationswhich, while reducing particle size, can also alter proteinstructure. For example, ultrasound can act as a potentdenaturant (39, 40); membranes are particularly susceptibleand sonication destroys membrane-associated histocompati-bility antigens (41). Heating appears less critical here thanfree radical formation and streaming (42, 43). The Frenchpress (38) is also not innocuous and denatures soluble proteinsin its usual operating range (about 1000 kg/cM2) (40).Erythrocyte membranes appear even more sensitive andexposure to about 300 kg/CM2 produces leakiness to K+ andincreased fragility (44), while exposure to 1000 kg/cM2appears to alter electrical conductivity (45).Some use sodium dodecyl sulfate to generate a solution
"reference state" (33, 35), but at the concentrations used(about 1 mg/mg of membrane protein) Na dodecyl sulfateacts as a potent protein perturbant, generating the antiparallel,B-conformation with poly(L-lysine) (46), producing nuclearmagnetic resonance evidence which indicates unfolding of
TABLE 1. Predicted ellipticities, 0, of variously sized sphericalshells at selected wavelengths and Felicities
He- Wave- 0 at external radiusb (nm X 10-3)licity length(%)- (nm) O 0.05 0.1 1.0
100 197 44,300 57,040 59,800 66,050100 210 -32,400 -30,210 -28,040 -21,830100 220 -35,300 -33,310 -33,200 -30,90050 197 10,000 6,280 8,100 12,25050 210 -16,900 -17,000 -16,400 -14,42050 220 -15,400 -14,850 -14,900 -14,22020 197 -24, 700 -24,100 -22,800 -19,80020 210 - 7,600 - 9,100 - 9,500 - 9,96020 220 - 3,100 - 3,800 - 3,970 - 4,200
* Nonhelical peptide in unordered state.b Shell thickness = 7.5 nm; 0 in degrees *cm2.dmole-1.o Solution.
membrane proteins (47), and inactivating numerous mem-brane enzymes (48, 49).
Organic solvents (50) may also perturb protein structure.Thus, 2-chloroethanol drastically alters protein helicity (28,29) while pentanol extraction (51) and acidic solvents (52)generate a-structures in membrane proteins. The latter canbe detected by infrared spectroscopy, but few apply thistechnique to biomembranes. All in all, the spectra engenderedby the listed procedures would appear to be even moreambiguous than those of untreated membranes.
MATERIALS AND METHODS
For application of the Mie theory we adopted the approachdeveloped by Gordon (34) and Gordon and Holzwarth (33)and were greatly aided by Dr. Gordon's generous gift of hiscomputer program (SCATCO). We take each membraneparticle as an isotropic spherical shell, 7.5 nm thick surround-ing an isotropic core (35, 36). We focus on particle radii of 75and 3500 nm. We treat each particle as a discrete, independentscatterer and neglect multiple scattering. We give the solventwithin each particle, as well as the bulk medium, a fixedrefractive index ml = m3 = 1.4, corresponding to water near200 nm. Dispersions of the refractive indices are neglected.We assumed a relative real refractive index of 1.2 for themembrane shell (27).
TABLE 2. Molecular weight and changes in optical rotatorydispersion of galactose-4-epimerase with variations of
ionic strength and temperature
Solvent (mI')(1 M DegreesTris- ORD cm2.HCl, Temp. Molecular trough deci-pH 7.5) 0C weight 820,wO (nm) mole-'
0.1 25 240,000 10. 0, 6.3 230 -0.01 25 120,000 6.3 230 42000.001 9 60,000 2.8,4.3 233 45400.001 25 60,000 2.8,4.3 236 4000
- 0.001 40 - 237 3700
from ref. 57.
Proc. Nat. Acad. Sci. USA 70 (1973)

3238 Biochemistry: Wallach et al.
We computed the complex refractive indices, M2R and M2L,of polylysine shells for right (R)- and left (L)-circularlypolarized light at a specific vacuum wavelength, X, of irradiat-ing light from established solution values of absorbance (53),CD (54), and ORD (55) of poly(ilysine). We used an RCA70/46 computer and Fortran IV compiled programs. Data arepresented in degrees * cm2 - dmol-'.
Experimental ORD and CD measurements were done on aCary model 6001 spectropolarimeter, calibrated with d-10-camphorsulfonic acid at 290 nm and using thermostatted1.000-, 0.500-, and 0.100-mm cells (Optical Cell Co. Inc.,Beltsville, Mid.). Measurements were at 260, except in thetemperature series. Baselines were run immediately after eachspectrum, with the same cell. ORD and CD spectra wereobtained on the same samples.Erythrocyte ghosts, normally oriented and inverted erythro-
cvte membrane vesicles, were prepared as described (56) andexamined on the same day. We used 7 mM phosphate (pH7.4) and 0.1 mg protein per ml for optical activity studies.Mean residue ellipticity [0] and rotation [m] assume a meanresidue weight of 130 (27).We equilibrated UIDP galactose-4-epimerase, purified from
Candida pseudo-tropicalis (4), with various buffers by gel-permeation chromatography on Sephadex G-50 (PharmaciaFine Chemicals).
Results and Discussion. We designed our studies to deter-mine (i) which aspects of membrane CD and ORD mightarise exclusively from turbidity and (ii) which features reflectthe inadequacy of synthetic polypeptides as general confor-mational standards. The CD, ORD, and absorbance dataavailable for poly(L-lysine) are sufficient to test these basicquestions.Membrane densities generally lie closer to 1.18 gl/'cm3 (58)
than the shell density of 1.5 assumed by Gordon and Holz-warth (33). However, calculations at 197, 210, and 220 nm,for a-helix: unordered proportions of 1.00, 0.5, and 0.2, showthat density variations from 1.15-1.50 do not change thecomputed ellipticities of 75- and 3500-nm particles by morethan 1%.We use a relative refractive index, n = 1.2 (33, 34). This
matches our experimental data (30). We further neglectdispersive effects on n and assume a single refractive indexvalue for the entire membrane shell. If the membranes com-prise separated lipid and protein patches _ than the wave-length of light, this assumption would introduce error, butthis problem does not apply to the 75-nm particles.Our Mie-scattering computations for poly(L-lysine) at [a-
helix:unordered] proportions from 1.00-0.20 clearly demon-strate that light-scattering distortions of CD are strongly con-formation-sensitive (Figs. 1 and 2). This is most marked below215 nm, largely because of the strong negative CD of "un-ordered" polypeptide near 200 nm. The distortions varystrongly with particle radius, conformation, and wavelength(Table 1). The computed CD of hypothetical 75-nm particlesfollows the solution spectrum closely between 190 and 230nm. However, above 230 nm even such small vesicles intro-duce a turbidity artifact, broadening the "222-nm" CD bandsufficiently to produce a shift of the ORD minimum from 233to235-236nmOur computations clearly demonstrate that one cannot
predict or correct light-scattering distortions unambiguously
this is exactly what one wishes to determine by optical activitymeasurements.Turning now to membrane CD (Figs. 3 and 4), we note
that one can obtain a "good fit" near 220 nm, knowing particlesize, assuming suitable conformational proportions, andapplying the Mie corrections. However, a fit near 220 nminvariably produces a major mismatch below 215 nm. Thisdilemma has been previously encountered with small, well-characterized, globular proteins (21, 24-26) and is not uniqueto membranes.We suspect that the discrepancies can be qualitatively
explained as follows (a) globular and membrane proteins tendto give low "helical" signals (near 222, 210, and 195 nm) forreasons given in the introduction, e.g., short helical segments,distortions from perfect helicity, and apolar environments;(b) in native proteins, peptide linkages not in a-helical or f3-structured disposition, comprise diverse, irregular, nonex-tended, but nonrandom arrays; the summed CD contributionof such segments is likely to be diffuse. Because of (a) and (b),reference to synthetic polypeptide standards tends to under-estimate overall "helicity" and exaggerate the proportion of"unordered" peptide segments. Accordingly, we agree thatmembrane proteins may contain more helix than indicated bytheir optical activity. However, no spectroscopic technique inhand can quantify "helicity" unambiguously. For example thelesser CD amplitude of Ehrlich ascites carcinoma plasmamembranes, compared with erythrocyte ghosts, could reflectgreater overall "helicity" in the latter, or a greater proportionof long helical segments.
In general, the CD band near 222 nm is so broad that itsposition is difficult to assign precisely [even in purely helicalpoly(rlysine) (48)]. However, scattering effects can accountfor small red shifts of this minimum (Figs. 1 and 2), particu-larly at high helicity. Moreover, they definitely broaden the"222-nm band" to the red sufficiently and can thus account forthe long-observed "red shift" of the ORD minimum of membranes.
Since scatter-corrected spectra match observed membraneCD poorly in the 7r-r* regions, we wonder about their reli-ability in other spectral regions, particularly because of theuncertainties in the optical activities of f-structured peptidesand the behavior exemplified by galactose-4-epimerase. Thisoligomeric protein yields infrared spectra, which signal thepresence of considerable p-structured peptide (4). Importantly,at low ionic strength the protein dissociates into presumptivemonomers, as seen upon analytical ultracentrifugation (Table2), and separation into smaller subunits is associated with anORD red shift of 6.0-7.0 nm. This shift is clearly due tobroadening of the "222-nm" band, and might reflect a changein the proportion and/or type of ,-structure (12). An ORD"red shift," thus, cannot be considered unique to membranesand be unambiguously attributed to light scattering.The Mie computations can, however, explain why the ORD
minimum does not shift after erythrocyte ghosts are frag-mented into small vesicles by a nondenaturing process. Thus,our erythrocyte ghosts exhibited the optical activity reportedbefore (27), with the ORD trough at 236 nm ([m] = -6150),the ORD crossover at 226.5 nm, and the first CD minimumat 223.2 nm ([0] = -13,800). Endo- or exocytotic disruptionof the ghosts into vesicles with diameters of 100 nm (56)reduces light scattering but does not measurably influence theoptical activity parameters under study. The above observa-tions can be explained by the fact, illustrated in all figures,
Proc. Nat. Acad. Sci. USA 70 (1973)
without knowing the conformational proportionof the sample; but

Proc. Nat. Acad. Sci. USA 70 (1973)
that the scattering-induced widening of the "222-nm" CDband changes negligibly as vesicle radius is decreased from3500 to 75 nm. Our computations also show that comminutionof membranes to small vesicles [e.g., sonication of liver "bilefronts" (50)] without alteration of protein structure cannot beexpected to "correct" membrane optical activity in an unam-
biguous fashion.It appears that the red shift of the "helical" ORD trough
can arise wholly, but not uniquely, as a turbidity effect.However, the other features of membrane optical activitycannot be attributed solely to light scattering. Rather, thematter of interpreting membrane optical activity is, in general,indistinguishable from that of using CD and ORD measure-
ments for the conformational analysis of globular proteins.
We thank Dr. David Gordon for the SCATCO program, Dr.T. L. Steck for the erythrocyte membrane preparations, andDr. H. Kalckar for use of the spectropolarimeter. Supported byGrants PRA-78 from the American Cancer Society (D.F.H.W.),GB 23123 from the National Science Foundation, CA 12178 fromthe United States Public Health Service, AT(30-1)-3748 from theAtomic Energy Commission, and HS 00696-02 from the UnitedStates Public Health Service.
1. Blake, C. C. F., Koenig, D. F., Mair, G. A., North, A. C. T.& Sarma, V. R. (1965) Nature 206, 757-761.
2. Birktoft, J. J., Blow, D. M., Henderson, R. & Steitz, T. A.(1970) Phil. Trans. Roy. Soc. London, Ser. B. 257, 67-76.
3. Phillips, D. C. (1967) Proc. Nat. Acad. Sci. USA 57, 484-495.
4. Bertland, A. U. & Kalckar, H. M. (1968) Proc. Nat. Acad.Sci. USA 61, 629-635.
5. Graham, J. M. & Wallach, D. F. H. (1969) Biochim. Bio-phys. Acta 193, 225-227.
6. Graham, J. M. & Wallach, D. F. H. (1971) Biochim. Bio-phys. Acta 241, 180-194.
7. Avruch, J. & Wallach, D. F. H. (1971) Biochim. Biophys.Acta 241, 249-253.
8. Choules, G. L. & Bjorklund, R. (1970) Biochemistry 9,4759-4767.
9. Tinoco, E., Jr., Woody, P. W. & Bradley, D. F. (1963) J.Chem. Phys. 38, 1317-1325.
10. Bradley, D. F., Tinoco, I., Jr. & Woody, R. W. (1963)Biopolymers 1, 239-267.
11. Yaron, A., Katchalski, E., Berger, A., Fasman, G. D. &Sober, H. A. (1971) Biopolymers 10, 1107-1120.
12. Perutz, M. F., Muirhead, H., Cox, J. M. & Goaman, L. C.(1968) Nature 219, 131-139.
13. Carver, J. P., Shechter, E. & Blout, E. R. (1966) J. Amer.Chem. Soc. 88, 2550-2561.
14. Stevens, L., Townend, R., Timasheff, S., Fasman, G. &Potter, J. (1968) Biochemistry 7, 3717-3720.
15. Tooney, N. M. & Fasman, G. D. (1968) J. Mol. Biol. 36,355-369.
16. Tiffany, M. L. & Krimm, S. (1969) Biopolymers 8, 347-359.17. Fasman, G. D., Hoving, H. & Timasheff, S. N. (1970)
Biochemistry 9, 3316-3324.18. Dearborn, D. G. & Wetlaufer, D. B. (1970) Biochem. Bio-
phys. Res. Commun. 39, 314-320.19. Magar, M. E. (1971) J. Theor. Biol. 33, 105-119.20. Stryer, L. (1965) J. Mol. Biol. 13, 482-495.21. Straus, J. H., Gordon, A. S. & Wallach, D. F. H. (1969)
Eur. J. Biochem. 11, 201-212.
22. Karlson, R. H., Norland, V. S., Fasman, G. D. & Blout,E. R. (1960) J. Amer. Chem. Soc. 82, 2268-2275.
23. Malcolm, B. R. (1970) Biopolymers 9, 911-922.24. Saxena, V. P. & Wetlaufer, D. B. (1971) Proc. Nat. Acad.
Sci. USA 68, 969-972.25. Rosenkranz, H. & Scholtan, W. (1971) Hoppe Seyler's Z.
Physiol. Chem. 352, 896-904.26. Chen, Y.-H., Yang, J. T. & Martinez, H. M. (1972) Bio-
chemistry 11, 4120-4131.27. Gordon, A. S., Wallach, D. F. H. & Straus, J. H. (1969)
Biochim. Biophys. Acta 183, 405-416.28. Lenard, J. & Singer, S. J. (1966) Proc. Nat. Acad. Sci. USA
56, 1828-1835.29. Wallach, D. F. H. & Zahler, P. H. (1966) Proc. Nat. Acad.
Sci. USA 56, 1552-1559.30. Wallach, D. F. H., Kamat, V. B. & Gail, M. H. (1966) J.
Cell Biol. 30, 601-621.31. Dickerson, R. E., Kopka, M. L., Wienzierl, J. E., Eisenberg,
D. & Margoliash, E. (1967) 154th National Meeting of theAmerican Chemical Soc.
32. Schneider, A. S. (1971) Chem. Phys. Lett. 8, 604-608.33. Gordon, D. J. & Holzwarth, G. (1971) Proc. Nat. Acad.
Sci. USA 68, 2365-2369.34. Gordon, D. J. (1972) Biochemistry 11, 413-420.35. Urry, E. W. (1972) Biochim. Biophys. Acta 265, 115-168.36. Gordon, D. J. & Holzwarth, G. (1971) Arch. Biochem.
Biophys. 142, 481-488.37. Schneider, A. S., Schneider, M. J. T. & Rosenheck, K. (1970)
Proc. Nat. Acad. Sci. USA 66, 793-798.38. Glaser, M. & Singer, S. J. (1971) Biochemistry 10, 1780-
1787.39. Weissler, A. J. (1960) J. Acoust. Soc. Amer. 32, 1208-1212.40. Joly, M. (1965) A Physicochemical Approach to the Dena-
turation of Proteins (Academic Press, New York), pp. 9-16.41. Haughton, G. (1966) Transplantation 4, 238-244.42. Grabar, P. (1953) Adv. Biol. Med. Phys. 3, 191-237.43. Hughes, D. E. (1965) in Ultrasonic Energy, ed. Kelly, E.
(University of Illinois Press, Urbana, II.), pp. 9-22.44. Yamoto, H. (1952) Oyakama Igakkai Zasshi64, 874-876.45. Mitayake, T. (1957) Oyakama Igakkai Zasshi 69, 461-463.46. Sarkar, P. K. & Doty, P. (1966) Proc. Nat. Acad. Sci. USA
55, 981-989.47. Chapman, D., Kamat, V. B., DeGier, J. & Penckett, S. A.
(1968) J. Mol. Biol. 31, 101-114.48. Wallach, D. F. H. (1969) J. Gen. Physiol. 54, 3'-268.49. Dunnick, J. K., Marinetti, G. V. & Greenland, P. (1972)
Biochim. Biophys. Acta 266, 684-694.50. Masotti, L., Urry, D. W., Krivacic, J. R. & Long, M. M.
(1972) Biochim. Biophys. Acta 266, 7-17.51. Singer, J. A. & Morrison, M. (1972) Biochim. Biophys.
Acta 274, 64-70.52. Wallach, D. F. H. & Zahler, P. H. (1968) Biochim. Biophys.
Acta 150, 186-193.53. McDiarmid, R. (1965) Ph.D. Thesis, Harvard University,
Cambridge, Massachusetts.54. Greenfield, N. & Fasman, G. D. (1969) Biochemistry 8,
4108-4116.55. Greenfield, N. & Fasman, G. D. (1967) Biochemistry 6,
1630-1637.56. Steck, T. L., Weinstein, R. S., Straus, J. H. & Wallach,
D. F. H. (1970) Science 168, 255-257.57. Bertland, L. H. & Bertland, A. U. (1971) Biochemistry 10,
3145-3150.58. Wallach, D. F. H. & Kamat, V. B. (1964) Proc. Nat. Acad.
Sci. USA 52, 721-728.
Membrane Optical Activity 3239
Related Documents











