Induction, distribution and modulation of upper airway allergic inflammation in mice I. HUSSAIN*, D. RANDOLPH*, S. L. BRODY², S.-K. SONG‡, A. HSU*, A. M. KAHN*, D. D. CHAPLIN*,§ and D. L. HAMILOS* *Division of Allergy and Immunology, ²Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine, ‡Department of Chemistry at Washington University School of Medicine, and §Howard Hughes Medical Institute, St Louis, USA Summary Background To further elucidate mechanisms of human allergic rhinosinusitis, we studied the induction, distribution and modulation of allergen-induced upper airway inflammation in a BALB/c mouse model. Methods Allergic inflammation induced with ovalbumin (OVA) by intraperitoneal (IP) injection in alum was compared to repeated intranasal instillation. The type and distribution of inflammatory cells was compared in the respiratory and olfactory epithelial compartments. Eosinophil distribution was assessed using Scarlet Red stain and a polyclonal antibody recognizing eosinophil major basic protein (MBP). The role of interleukin (IL)-5 in upper airway inflammation was tested by administration of polyclonal anti-IL-5 antibody during the sensitization protocol. Results Unsensitized control mice receiving saline failed to develop upper airway eosinophil infiltration. IP OVA-sensitized mice developed marked upper airway mucosal eosinophil infiltration after aerosol OVA challenge, whereas repeated intranasal instillation of OVA produced qualitatively similar, but less intense eosinophil infiltration. Using either sensitization protocol, eosinophil infiltration was seen in areas of the lower portion of the nasal septum, the floor and the lower lateral walls of the mid-caudal region of the nasal cavity. Immunofluorescence staining for MBP confirmed this distribution of eosinophils but also demonstrated some eosinophils in the maxillary sinuses and in circumscribed regions of the ethmoturbinates. All areas of eosinophil infiltration were lined by respiratory epithelium. The selective infiltration of respiratory but not olfactory epithelium by eosinophils was unassociated with a measurable induction of epithelial ICAM-1 or eotaxin expression. OVA-induced upper airway eosinophil infiltration was found to be IL-5 dependent, since administration of a polyclonal anti-IL-5 antibody (TRFK-5) during OVA sensitization resulted in a marked modulation (80% decrease) in eosinophil infiltration in response to subsequent OVA challenge. Conclusion The mouse upper airway, specifically in areas containing respiratory epithelium, is a target for OVA-induced allergic inflammation. This selective infiltration of respiratory, but not olfactory, epithelium is, in part, dependent upon IL-5. This model is useful for further dissection of the inflammatory response with genetic manipulations and targeted immunological approaches. Keywords: allergic, inflammation, ovalbumin, eosinophil, mice, respiratory, olfactory Clinical and Experimental Allergy, Vol. 31, pp. 1048–1059. Submitted 29 September 2000; revised 21 December 2000; accepted 22 January 2001. Clinical and Experimental Allergy, 2001, Volume 31, pages 1048–1059 1048 q 2001 Blackwell Science Ltd Correspondence: Dr Daniel L. Hamilos, Washington University School of Medicine, Division of Allergy and Immunology, Box 8122, 660 South Euclid Avenue, St Louis, MO 63110, USA

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Induction, distribution and modulation of upper airway
allergic inflammation in mice
I. HUSSAIN*, D. RANDOLPH*, S. L. BRODY², S.-K. SONG³, A. HSU*,A. M. KAHN*, D. D. CHAPLIN*,§ and D. L. HAMILOS*
*Division of Allergy and Immunology, ²Division of Pulmonary and Critical Care Medicine, Department of Internal
Medicine, ³Department of Chemistry at Washington University School of Medicine, and §Howard Hughes Medical
Institute, St Louis, USA
Summary
Background To further elucidate mechanisms of human allergic rhinosinusitis, we
studied the induction, distribution and modulation of allergen-induced upper airway
inflammation in a BALB/c mouse model.
Methods Allergic inflammation induced with ovalbumin (OVA) by intraperitoneal (IP)
injection in alum was compared to repeated intranasal instillation. The type and
distribution of inflammatory cells was compared in the respiratory and olfactory epithelial
compartments. Eosinophil distribution was assessed using Scarlet Red stain and a
polyclonal antibody recognizing eosinophil major basic protein (MBP). The role of
interleukin (IL)-5 in upper airway inflammation was tested by administration of polyclonal
anti-IL-5 antibody during the sensitization protocol.
Results Unsensitized control mice receiving saline failed to develop upper airway
eosinophil infiltration. IP OVA-sensitized mice developed marked upper airway mucosal
eosinophil infiltration after aerosol OVA challenge, whereas repeated intranasal instillation
of OVA produced qualitatively similar, but less intense eosinophil infiltration. Using either
sensitization protocol, eosinophil infiltration was seen in areas of the lower portion of the
nasal septum, the floor and the lower lateral walls of the mid-caudal region of the nasal
cavity. Immunofluorescence staining for MBP confirmed this distribution of eosinophils
but also demonstrated some eosinophils in the maxillary sinuses and in circumscribed
regions of the ethmoturbinates. All areas of eosinophil infiltration were lined by respiratory
epithelium. The selective infiltration of respiratory but not olfactory epithelium by
eosinophils was unassociated with a measurable induction of epithelial ICAM-1 or eotaxin
expression. OVA-induced upper airway eosinophil infiltration was found to be IL-5
dependent, since administration of a polyclonal anti-IL-5 antibody (TRFK-5) during OVA
sensitization resulted in a marked modulation (80% decrease) in eosinophil infiltration in
response to subsequent OVA challenge.
Conclusion The mouse upper airway, specifically in areas containing respiratory
epithelium, is a target for OVA-induced allergic inflammation. This selective infiltration
of respiratory, but not olfactory, epithelium is, in part, dependent upon IL-5. This model is
useful for further dissection of the inflammatory response with genetic manipulations and
targeted immunological approaches.
Keywords: allergic, inflammation, ovalbumin, eosinophil, mice, respiratory, olfactory
Clinical and Experimental Allergy, Vol. 31, pp. 1048±1059. Submitted 29 September
2000; revised 21 December 2000; accepted 22 January 2001.
Clinical and Experimental Allergy, 2001, Volume 31, pages 1048±1059
1048 q 2001 Blackwell Science Ltd
Correspondence: Dr Daniel L. Hamilos, Washington University School of
Medicine, Division of Allergy and Immunology, Box 8122, 660 South
Euclid Avenue, St Louis, MO 63110, USA

Introduction
Allergic rhinitis is a common chronic illness, affecting
approximately 10% of the population in the United States
of America [1]. Natural allergen exposure is associated
with chronic inflammation and mucosal oedema of the
respiratory epithelium in nasal and sinus passages giving
rise to symptoms of nasal congestion, rhinorrhea, nasal
blockage and reduced olfaction [2,3]. These symptoms
have a significant negative impact on quality of life, and
account for billions of dollars of health care expenditure
yearly on anti-allergic medications [4].
Although numerous studies have examined the induction
of allergic inflammation in the lower airways, there have
been relatively few such studies in the upper airway. While
the upper and lower airway mucosae are similar, there are
differences in innervation, connective tissue elements,
submucosal lymphatic structures and specialized epithelia
(such as the olfactory epithelium) that may affect the
quality or intensity of the inflammatory response. It is
likely that eosinophils play a major role in the allergic
response in both the upper and lower airways [5±7], and
studies in humans and rodents suggest a role for epithelial
induction of ICAM-1 and eotaxin in promoting eosinophil
infiltration [8±13]. Furthermore, the upper airway response
may differ from the lower airways in terms of its response
to locally applied allergen. Hence, there is a need for
experimental systems that can be easily modulated to
examine localization and dynamics of the allergic inflam-
mation in the upper airway after either systemic vs.
intranasal sensitization.
Similarly, the target epithelium for allergic inflamma-
tion is unclear. Human allergic rhinitis is associated with
a reduced olfactory function during seasonal allergen
exposure but not out of season [14±17]. This question is
difficult to address in humans but can be addressed in
mice, since the olfactory epithelium occupies a much
larger fraction of the upper airway area and is easily
identified by its anatomic localization and expression of
an epithelial cell-specific marker, olfactory marker protein
(OMP) [18].
In the study of the pathophysiology of asthma, several
groups have employed a model of intraperitoneal (IP)
sensitization followed by aerosol challenge with chicken
ovalbumin (OVA) to induce allergic inflammation in the
lower airways [19±26]. In this model, there is a
characteristic eosinophilic inflammatory response that is
dependent upon elaboration of Th2-type cytokines, parti-
cularly IL-5 [25±27]. Interleukin-5 is known to increase the
survival of eosinophils in tissues and to promote eotaxin-
induced airway eosinophil infiltration [28]. Because the
respiratory epithelial cells and inflammatory responses of
the upper airway share many characteristics with the lower
airway, we also wished to determine whether OVA-induced
upper airway allergic inflammation is IL-5 dependent.
Materials and methods
OVA sensitization and challenge protocols
Female BALB/c mice 8±12 weeks of age (Harlan Labora-
tories, Indianapolis, IN, USA) were housed in a pathogen±
free barrier facility and maintained on OVA-free diets. All
experimental animals used in this study were under a
protocol approved by the Animal Studies Committee of
Washington University School of Medicine.
Mice were sensitized with OVA given intraperitoneally
(IP) or intranasally (see Fig. 1). In the IP protocol, each
mouse received 8 mg of OVA (Chicken ovalbumin, Grade
V, Sigma, St Louis, MO, USA) adsorbed to 2 mg of alum
(Sigma) in 0.5 mL of sterile phosphate buffered saline
(PBS) on day 1, with a booster on day 6. Age-matched
control mice received only alum 2 mg in 0.5 cc of PBS on
the same days. OVA-sensitized mice received 1% OVA
(wt/vol) by aerosol on day 13 and 19 in a small plexiglas
chamber device attached to a clinical nebulizer (Ultra-Neb
99, DeVilbiss Health Care, Somerset, PA, USA). On each
challenge day, mice received aerosolized OVA for 20 min,
first in the morning and then in the afternoon. Mice were
killed 24±72 h after the last challenge. In the intranasal
sensitization protocol, mice received 10 mL of 1% OVA
intranasally to each nostril for 10 consecutive days. Sham-
sensitized mice received 10 mL of saline intranasally to
each nostril on the same days. Mice were killed 48 h after
the last intranasal dose.
In separate experiments, mice were treated with 100 mg of
rat antimurine IL-5 monoclonal antibody TRFK-5 (ATCC,
Manassas, VA, USA) administered IP on days 4, 6 and 8 during
the intranasal OVA sensitization protocol as previously
described [25]. Control mice received 100 mg of isotype-
matched rat IgG IP on the same days. Mice were killed 48 h
following 10 consecutive days of intranasal OVA sensitization.
Mouse sinus MRI
Upper airway anatomy was defined by high resolution magnetic
resonance imaging (MRI) scans. Mice were anaesthetized with
halothane/O2 mixture (5% induction, 1.5% maintenance). A
1.5-cm outer diameter circular surface coil was placed on the
mouse head between the rostral aspect of the snout and the eyes
for MR signal reception while a 12-cm inner diameter
Helmholtz coil was used as the transmitter. A multislice spin
echo imaging sequence was used for the data collection. The
imaging parameters used were: repetition time 2 s, echo time
18 msec, field of view 1.5 cm2, data matrix 256 � 256,
in-plane resolution 60 mm, and slice thickness 500 mm.
Upper airway allergic inflammation in mice 1049
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059

Upper airway sectioning, staining and immunohistochem-
istry
Mice were anaesthetized with ketamine/xylazine then killed
by cardiac exsanguination. Blood was flushed from the
circulation by intracardiac injection of sterile Dulbecco's
PBS. Heads were separated from the body, and the scalp was
removed. Whole heads were fixed in 10% buffered formalin
for 24±48 h. After fixation, the heads were decalcified in
14% EDTA at 218C for 2 week. After decalcification, heads
were sectioned coronally into four regions for paraffin
embedding: A at the incisor teeth, C at the anterior margin of
orbit, B halfway between A and C, and D at the posterior
margin of the orbit [29±31]. Tissue sections were cut at
5 mm thickness for evaluation.
To identify epithelial cells, tissue sections were deparaffi-
nized and hydrated and then incubated with a rat anti-mouse
Pan-cytokeratin (BD PharMingen, SanDiego, CA, USA)
1 : 200 diluted in Saponin buffer (0.5% Saponin in PBS) with
3% horse serum overnight at 4 8C. Control slides were
incubated with isotype-matched rat antibody under similar
conditions. To identify respiratory epithelial cells, antibody
directed toward HFH-4 was used. HFH-4 is a transcription
factor with tissue-specific expression on epithelial cells
possessing motor but not sensory cilia [32]. Rabbit anti-rat
HFH-4 polyclonal antibody (diluted 1 : 500) was used on
paraformaldehyde-fixed samples in paraffin as previously
described [32]. To identify neuro-olfactory epithelial cells,
goat anti-serum against mouse olfactory marker protein (OMP,
dilution 1: 4000) (a generous gift of Dr Frank Margolis,
Baltimore, MD, USA) was used [18,33]. To identify epithelial
expression of ICAM-1, a hampster anti-mouse monoclonal
anti-ICAM-1 antibody was used at a dilution of 1 : 500 (3E2,
BD PharMingen). To identify epithelial expression of eotaxin,
a polyclonal goat anti-mouse eotaxin antibody was used (AF-
420-NA, R & D Systems, Minneapolis, MN, USA). A
polyclonal rabbit anti-mouse anti-serum for major basic
protein (a generous gift of Dr Gerald Gleich, Mayo Clinic,
Rochester, MN, USA) was used at a dilution of 1 : 500.
Secondary antibodies of the appropriate species labelled with
Fig. 1. OVA sensitization and challenge protocols. (a) IP sensitization protocol. (b) Intranasal sensitization protocol. (c) Intranasal
sensitization protocol combined with IP anti-IL-5 protocol. See Materials and methods for details of each protocol.
1050 I. Hussain et al.
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059

alkaline phosphatase or horseradish peroxidase (Vector
Laboratories, Inc., Burlington, CA, USA) was used for
detection, and sections were counterstained with haema-
toxylin. For immunofluorescence, antibody binding was
visualized using a fluorescent CY3-labelled anti-rabbit IgG
secondary antibody. Control experiments were done with
isotype-specific IgG for each antibody.
Biebrich Scarlet staining for eosinophils
To detect basic protein at alkaline pH (pH � 9.5), Biebrich
Scarlet stain, previously shown to be highly specific for
eosinophils, was used [34,35]. tissue sections were
deparaffinized, rehydrated gradually to 30% ethanol and
washed in PBS for 5 min. Slides were immersed in
Biebrich Scarlet stain for 1 h at 218C. Biebrich Scarlet
stain was prepared by mixing 1.9 gram borax with 20 mg
of Biebrich Scarlet powder (Sigma) in 50 mL of water and
titrating the pH to 9.5. Slides were counterstained with
methyl green for 2 min, washed with 100% ethanol for one
min and dehydrated before applying coverslips.
Cell quantification
The number of eosinophils was quantified per unit of
epithelial basement membrane to a uniform depth of
submucosa encompassing an area of 1.26 mm2 using an
eyepiece reticule. The average of cells counted in at least 6
sections was used for data analysis. All slides were
examined by a single observer, blinded to the experimental
condition. The percentage reduction in numbers of upper
airway eosinophils caused by anti-IL-5 antibody treatment
was calculated using the formula:
% reduction � ��OVA-induced eosinophils�/�OVA
1 anti-IL-5Ab-induced eosinophils��4 ��OVA-induced eosinophils�2 �Saline-induced eosinophils��
Data analysis
Data from each experimental condition were averaged
and compared by non-parametric Mann±Whitney U-test
[36]. A P-value of 0.05 was regarded as statistically
significant.
Results
Mouse upper airway anatomy
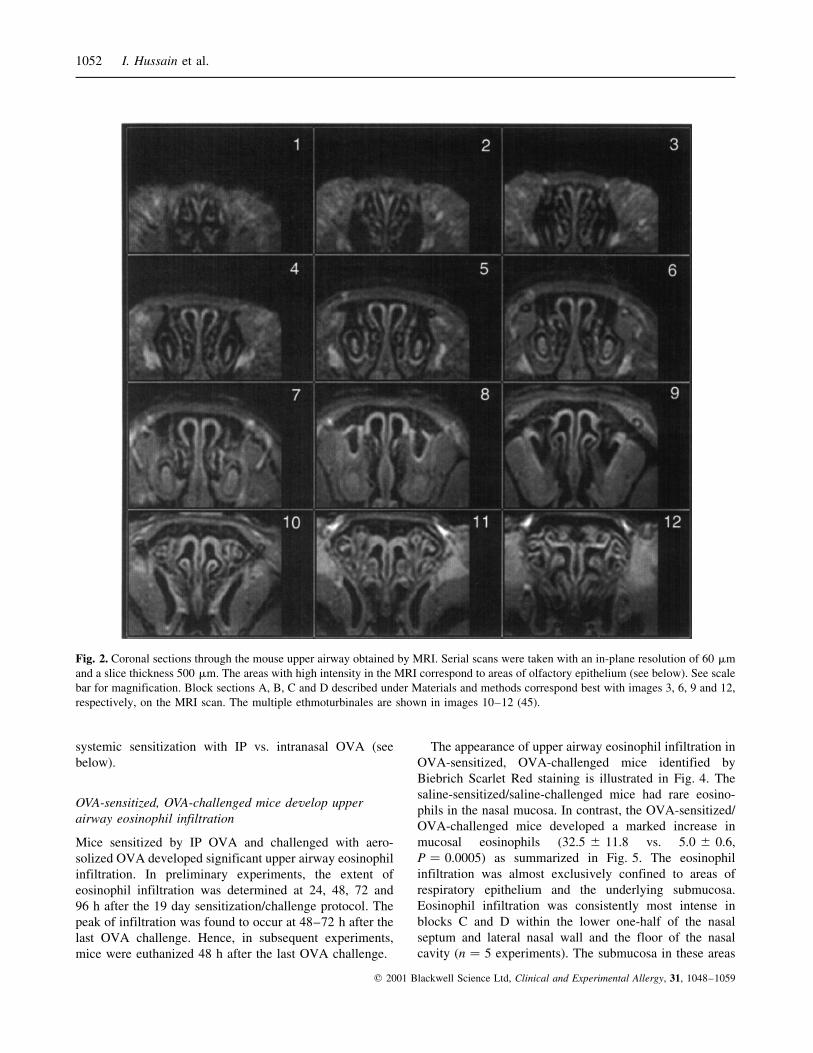
To define the distribution of inflammation in response to
OVA, the anatomy of the mouse upper airway needed to be
defined. For histological analysis, the upper airway was
sectioned rostral to caudal into four blocks A, B, C and D as
described under Materials and methods. Because the upper
airway anatomy is quite different in these four areas, we
illustrate the anatomy of the mouse upper airway in coronal
sections taken rostral to caudal by MRI scanning. The
images corresponding to the caudal ends of each block are
illustrated in Fig. 2. Key structures are identified in Fig. 3
(see below).
Distribution of respiratory and olfactory epithelium in the
mouse upper airway
The mouse olfactory epithelium produces a specific
protein known as olfactory marker protein or OMP
[18,33]. The BALB/c mouse olfactory epithelium was
found to have a characteristic high-intensity MRI signal
(shown in Fig. 2) and a thick, multinucleated appearance
on H & E stained tissues. The distribution of olfactory
epithelium was confirmed by immunostaining with OMP
anti-serum. OMP-positive epithelium was found in the
superior region of the nasal cavity, including the superior
aspect of the septum and the lateral nasal walls (Fig. 3).
The superior half of the vomeronasal body, an olfactory
organ, was also defined by strong OMP expression
[18,33].
The distribution of cytokeratin-positive epithelium in the
mouse upper airway was determined by immunostaining
with a rat anti-mouse pancytokeratin antibody (data not
shown). Positive immunostaining was observed throughout
the entire upper airway but was most intense in the lower
half of the septum, the lateral nasal wall and the floor of the
nasal cavity. Olfactory epithelium also showed weak
immunostaining for cytokeratin.
The distribution of respiratory epithelium in the mouse
upper airway was also examined by immunostaining for the
transcription factor HFH-4 (Fig. 3). Using a polyclonal
anti-HFH-4 antibody, the distribution of HFH-4 immuno-
staining matched the distribution of high-intensity cyto-
keratin immunostaining, namely involving the floor of the
nasal cavity, the inferior half of the lateral wall and the
septum and maxillary sinuses. As expected, HFH-4
immunostaining was specific for respiratory epithelium
and exclusive of cells that expressed OMP. In subsequent
experiments, we found that OVA-induced upper airway
eosinophil infiltration was specific for areas lined by
respiratory epithelium.
Compartments of bone marrow were also identified in
the superior portion of the nasal septum and just superior
to the nasal cavity within the skull (refer to Fig. 6 below).
The eosinophil density in this compartment was used to
assess the magnitude of inflammatory response to
Upper airway allergic inflammation in mice 1051
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059
systemic sensitization with IP vs. intranasal OVA (see
below).
OVA-sensitized, OVA-challenged mice develop upper
airway eosinophil infiltration
Mice sensitized by IP OVA and challenged with aero-
solized OVA developed significant upper airway eosinophil
infiltration. In preliminary experiments, the extent of
eosinophil infiltration was determined at 24, 48, 72 and
96 h after the 19 day sensitization/challenge protocol. The
peak of infiltration was found to occur at 48±72 h after the
last OVA challenge. Hence, in subsequent experiments,
mice were euthanized 48 h after the last OVA challenge.
The appearance of upper airway eosinophil infiltration in
OVA-sensitized, OVA-challenged mice identified by
Biebrich Scarlet Red staining is illustrated in Fig. 4. The
saline-sensitized/saline-challenged mice had rare eosino-
phils in the nasal mucosa. In contrast, the OVA-sensitized/
OVA-challenged mice developed a marked increase in
mucosal eosinophils (32.5 ^ 11.8 vs. 5.0 ^ 0.6,
P � 0.0005) as summarized in Fig. 5. The eosinophil
infiltration was almost exclusively confined to areas of
respiratory epithelium and the underlying submucosa.
Eosinophil infiltration was consistently most intense in
blocks C and D within the lower one-half of the nasal
septum and lateral nasal wall and the floor of the nasal
cavity (n � 5 experiments). The submucosa in these areas
Fig. 2. Coronal sections through the mouse upper airway obtained by MRI. Serial scans were taken with an in-plane resolution of 60 mm
and a slice thickness 500 mm. The areas with high intensity in the MRI correspond to areas of olfactory epithelium (see below). See scale
bar for magnification. Block sections A, B, C and D described under Materials and methods correspond best with images 3, 6, 9 and 12,
respectively, on the MRI scan. The multiple ethmoturbinales are shown in images 10±12 (45).
1052 I. Hussain et al.
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059
remained relatively thin, but there was a suggestion of
expansion in some areas with the most intense eosinophil
infiltration. In contrast, virtually no eosinophils were seen
in areas of olfactory epithelium.
Enhanced detection of OVA-induced upper airway eosino-
phil infiltration
Eosinophils were easily identified in areas of respiratory
epithelium by Scarlet Red staining. However, their identifica-
tion was less precise in the thicker olfactory epithelium. To
more precisely examine the entire mouse upper airway, we
used immunofluorescent staining with a primary MBP
antibody and a fluorescent CY3-conjugated secondary anti-
body and examined the distribution of eosinophils in OVA-
sensitized/OVA-challenged mice. Figure 6 shows a colour
enhanced reconstruction of the entire mouse upper airway
corresponding to block D stained for MBP. The background
colour was enhanced to demonstrate the anatomy. The normal
red-orange fluorescence of the CY3 has been colour enhanced
to yellow for better visualization of eosinophils. As shown, the
predominant distribution of eosinophils by MBP staining was
similar to that detected by Scarlet Red stain, namely involving
the lower one-half of the nasal septum and lateral nasal wall
and the floor of the nasal cavity. In addition, a few eosinophils
could be detected in the epithelium of the maxillary sinuses,
the inferior portions of the inferior turbinates and in the ventral
scroll and the base of the second ethmoturbinates. Eosinophils
were only seen in areas lined with respiratory epithelium.
Upper airway eosinophil infiltration requires OVA sensiti-
zation followed by aerosol OVA challenge
To determine whether upper airway eosinophil infiltration
required both systemic OVA sensitization and aerosol OVA
challenge, we compared eosinophil infiltration in IP saline-
sensitized/intranasal saline-challenged, IP OVA-sensitized/
intranasal saline-challenged, and IP OVA-sensitized/intranasal
OVA-challenged mice. The extent of eosinophil infiltration
was determined by MBP immunofluorescence. As shown in
Fig. 7, the IP OVA-sensitized/intranasal saline mice showed
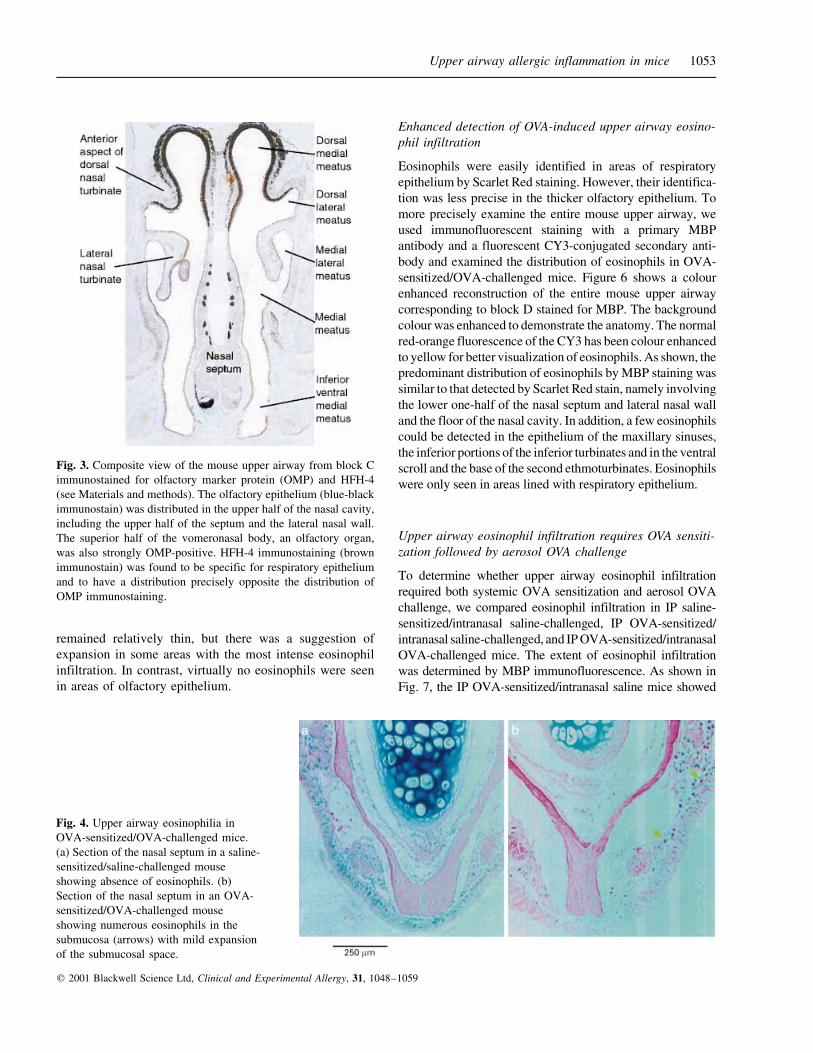
Fig. 4. Upper airway eosinophilia in
OVA-sensitized/OVA-challenged mice.
(a) Section of the nasal septum in a saline-
sensitized/saline-challenged mouse
showing absence of eosinophils. (b)
Section of the nasal septum in an OVA-
sensitized/OVA-challenged mouse
showing numerous eosinophils in the
submucosa (arrows) with mild expansion
of the submucosal space.
Fig. 3. Composite view of the mouse upper airway from block C
immunostained for olfactory marker protein (OMP) and HFH-4
(see Materials and methods). The olfactory epithelium (blue-black
immunostain) was distributed in the upper half of the nasal cavity,
including the upper half of the septum and the lateral nasal wall.
The superior half of the vomeronasal body, an olfactory organ,
was also strongly OMP-positive. HFH-4 immunostaining (brown
immunostain) was found to be specific for respiratory epithelium
and to have a distribution precisely opposite the distribution of
OMP immunostaining.
Upper airway allergic inflammation in mice 1053
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059
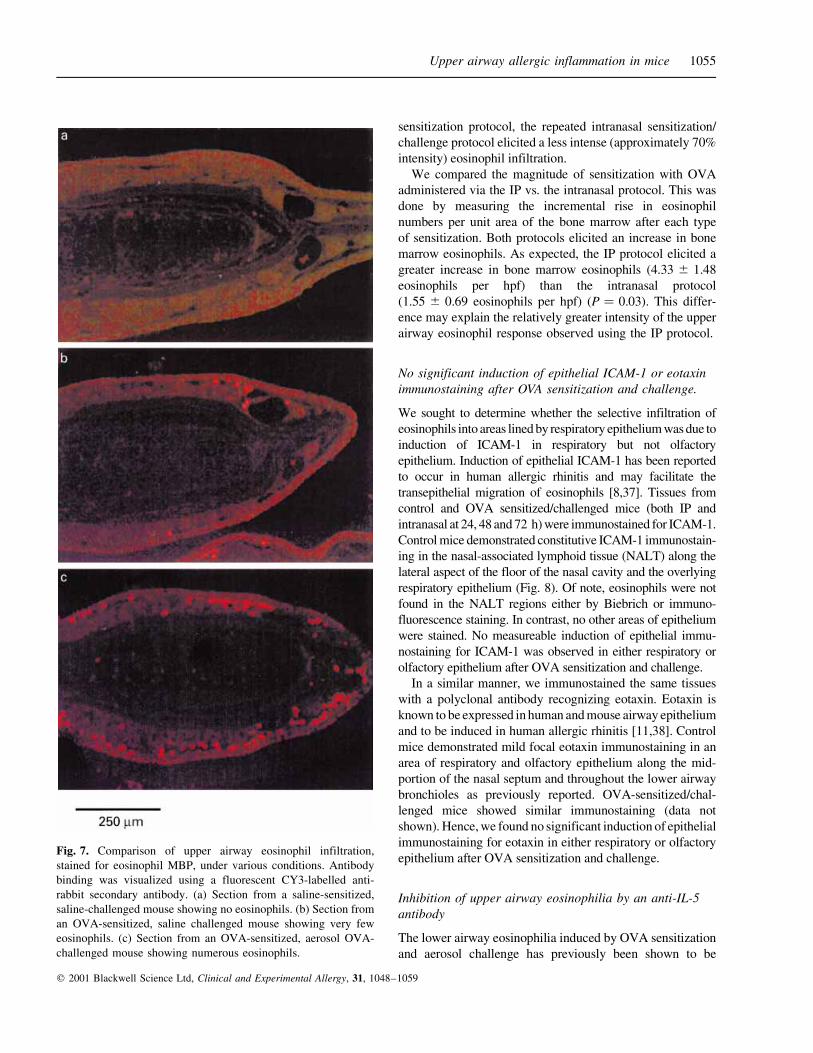
only rare upper airway eosinophils, similar to the IP saline-
sensitized/intranasal saline-challenged mice. In contrast, the IP
OVA-sensitized/intranasal OVA-challenged mice showed a
large increase in the numbers of eosinophils as expected.
Hence, IP OVA sensitization alone was insufficient to induce
upper airway eosinophil infiltration.
Intranasal vs. IP sensitization
To more closely approximate the process of intranasal
sensitization to allergens, BALB/c mice were sensitized by
intranasal instillation of 1% OVA (10 mL per nostril) daily for
10 consecutive days. This protocol was previously reported to
elicit a more selective IgE anti-OVA response, whereas the IP
OVA sensitization protocol elicited both an IgE and an IgG
anti-OVA response [25]. Control mice that received repeated
intranasal instillation of saline failed to develop upper airway
eosinophil infiltration. Mice administered repeated intranasal
OVA developed upper airway eosinophil infiltration in a
pattern similar to that observed after IP OVA sensitization and
aerosol OVA challenge. This infiltration was statistically
significant compared to intranasal saline alone (23.3 ^ 20.3
vs. 2.0 ^ 0.1, P � 0.02). In comparison with the IP
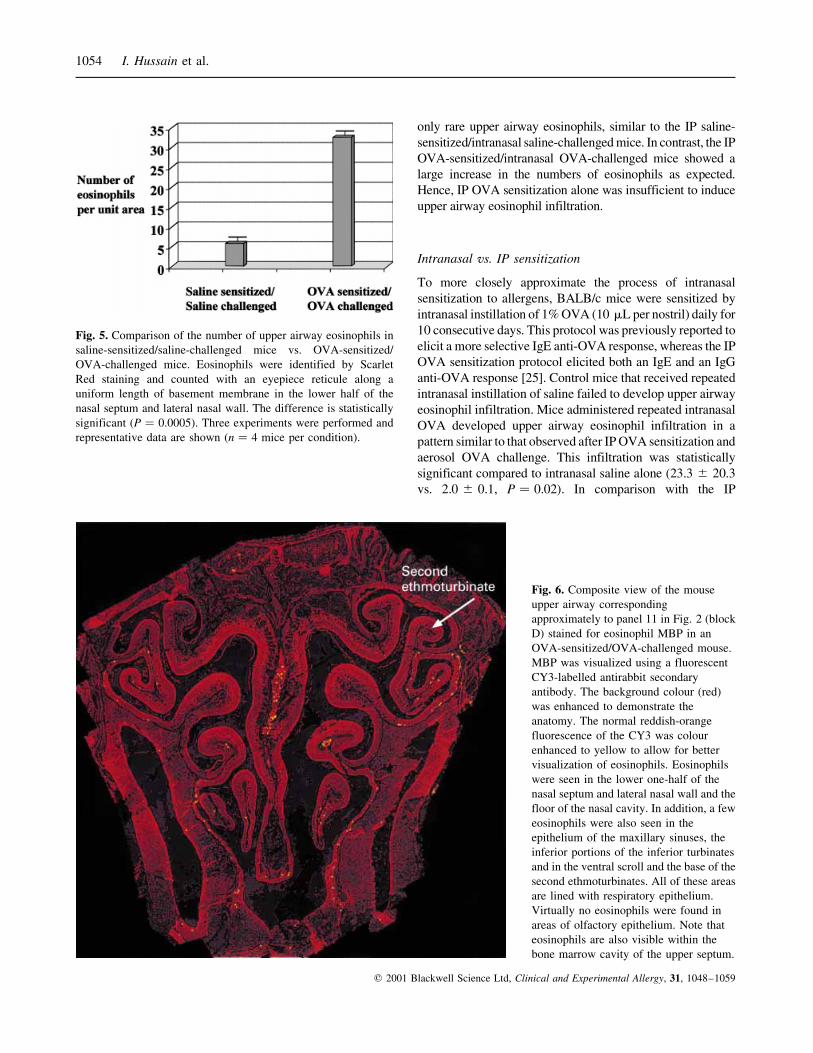
Fig. 6. Composite view of the mouse
upper airway corresponding
approximately to panel 11 in Fig. 2 (block
D) stained for eosinophil MBP in an
OVA-sensitized/OVA-challenged mouse.
MBP was visualized using a fluorescent
CY3-labelled antirabbit secondary
antibody. The background colour (red)
was enhanced to demonstrate the
anatomy. The normal reddish-orange
fluorescence of the CY3 was colour
enhanced to yellow to allow for better
visualization of eosinophils. Eosinophils
were seen in the lower one-half of the
nasal septum and lateral nasal wall and the
floor of the nasal cavity. In addition, a few
eosinophils were also seen in the
epithelium of the maxillary sinuses, the
inferior portions of the inferior turbinates
and in the ventral scroll and the base of the
second ethmoturbinates. All of these areas
are lined with respiratory epithelium.
Virtually no eosinophils were found in
areas of olfactory epithelium. Note that
eosinophils are also visible within the
bone marrow cavity of the upper septum.
Fig. 5. Comparison of the number of upper airway eosinophils in
saline-sensitized/saline-challenged mice vs. OVA-sensitized/
OVA-challenged mice. Eosinophils were identified by Scarlet
Red staining and counted with an eyepiece reticule along a
uniform length of basement membrane in the lower half of the
nasal septum and lateral nasal wall. The difference is statistically
significant (P � 0.0005). Three experiments were performed and
representative data are shown (n � 4 mice per condition).
1054 I. Hussain et al.
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059
sensitization protocol, the repeated intranasal sensitization/
challenge protocol elicited a less intense (approximately 70%
intensity) eosinophil infiltration.
We compared the magnitude of sensitization with OVA
administered via the IP vs. the intranasal protocol. This was
done by measuring the incremental rise in eosinophil
numbers per unit area of the bone marrow after each type
of sensitization. Both protocols elicited an increase in bone
marrow eosinophils. As expected, the IP protocol elicited a
greater increase in bone marrow eosinophils (4.33 ^ 1.48
eosinophils per hpf) than the intranasal protocol
(1.55 ^ 0.69 eosinophils per hpf) (P � 0.03). This differ-
ence may explain the relatively greater intensity of the upper
airway eosinophil response observed using the IP protocol.
No significant induction of epithelial ICAM-1 or eotaxin
immunostaining after OVA sensitization and challenge.
We sought to determine whether the selective infiltration of
eosinophils into areas lined by respiratory epithelium was due to
induction of ICAM-1 in respiratory but not olfactory
epithelium. Induction of epithelial ICAM-1 has been reported
to occur in human allergic rhinitis and may facilitate the
transepithelial migration of eosinophils [8,37]. Tissues from
control and OVA sensitized/challenged mice (both IP and
intranasal at 24, 48 and 72 h) were immunostained for ICAM-1.
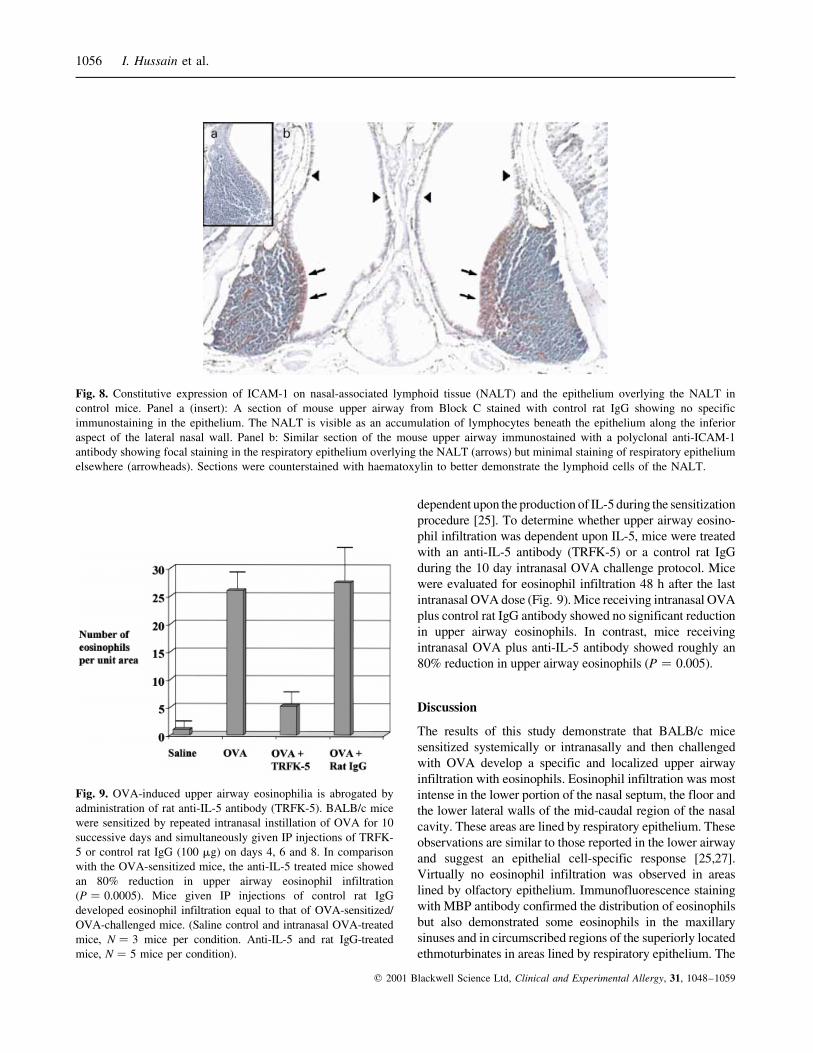
Control mice demonstrated constitutive ICAM-1 immunostain-
ing in the nasal-associated lymphoid tissue (NALT) along the
lateral aspect of the floor of the nasal cavity and the overlying
respiratory epithelium (Fig. 8). Of note, eosinophils were not
found in the NALT regions either by Biebrich or immuno-
fluorescence staining. In contrast, no other areas of epithelium
were stained. No measureable induction of epithelial immu-
nostaining for ICAM-1 was observed in either respiratory or
olfactory epithelium after OVA sensitization and challenge.
In a similar manner, we immunostained the same tissues
with a polyclonal antibody recognizing eotaxin. Eotaxin is
known to be expressed in human and mouse airway epithelium
and to be induced in human allergic rhinitis [11,38]. Control
mice demonstrated mild focal eotaxin immunostaining in an
area of respiratory and olfactory epithelium along the mid-
portion of the nasal septum and throughout the lower airway
bronchioles as previously reported. OVA-sensitized/chal-
lenged mice showed similar immunostaining (data not
shown). Hence, we found no significant induction of epithelial
immunostaining for eotaxin in either respiratory or olfactory
epithelium after OVA sensitization and challenge.
Inhibition of upper airway eosinophilia by an anti-IL-5
antibody
The lower airway eosinophilia induced by OVA sensitization
and aerosol challenge has previously been shown to be
Fig. 7. Comparison of upper airway eosinophil infiltration,
stained for eosinophil MBP, under various conditions. Antibody
binding was visualized using a fluorescent CY3-labelled anti-
rabbit secondary antibody. (a) Section from a saline-sensitized,
saline-challenged mouse showing no eosinophils. (b) Section from
an OVA-sensitized, saline challenged mouse showing very few
eosinophils. (c) Section from an OVA-sensitized, aerosol OVA-
challenged mouse showing numerous eosinophils.
Upper airway allergic inflammation in mice 1055
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059
dependent upon the production of IL-5 during the sensitization
procedure [25]. To determine whether upper airway eosino-
phil infiltration was dependent upon IL-5, mice were treated
with an anti-IL-5 antibody (TRFK-5) or a control rat IgG
during the 10 day intranasal OVA challenge protocol. Mice
were evaluated for eosinophil infiltration 48 h after the last
intranasal OVA dose (Fig. 9). Mice receiving intranasal OVA
plus control rat IgG antibody showed no significant reduction
in upper airway eosinophils. In contrast, mice receiving
intranasal OVA plus anti-IL-5 antibody showed roughly an
80% reduction in upper airway eosinophils (P � 0.005).
Discussion
The results of this study demonstrate that BALB/c mice
sensitized systemically or intranasally and then challenged
with OVA develop a specific and localized upper airway
infiltration with eosinophils. Eosinophil infiltration was most
intense in the lower portion of the nasal septum, the floor and
the lower lateral walls of the mid-caudal region of the nasal
cavity. These areas are lined by respiratory epithelium. These
observations are similar to those reported in the lower airway
and suggest an epithelial cell-specific response [25,27].
Virtually no eosinophil infiltration was observed in areas
lined by olfactory epithelium. Immunofluorescence staining
with MBP antibody confirmed the distribution of eosinophils
but also demonstrated some eosinophils in the maxillary
sinuses and in circumscribed regions of the superiorly located
ethmoturbinates in areas lined by respiratory epithelium. The
Fig. 8. Constitutive expression of ICAM-1 on nasal-associated lymphoid tissue (NALT) and the epithelium overlying the NALT in
control mice. Panel a (insert): A section of mouse upper airway from Block C stained with control rat IgG showing no specific
immunostaining in the epithelium. The NALT is visible as an accumulation of lymphocytes beneath the epithelium along the inferior
aspect of the lateral nasal wall. Panel b: Similar section of the mouse upper airway immunostained with a polyclonal anti-ICAM-1
antibody showing focal staining in the respiratory epithelium overlying the NALT (arrows) but minimal staining of respiratory epithelium
elsewhere (arrowheads). Sections were counterstained with haematoxylin to better demonstrate the lymphoid cells of the NALT.
Fig. 9. OVA-induced upper airway eosinophilia is abrogated by
administration of rat anti-IL-5 antibody (TRFK-5). BALB/c mice
were sensitized by repeated intranasal instillation of OVA for 10
successive days and simultaneously given IP injections of TRFK-
5 or control rat IgG (100 mg) on days 4, 6 and 8. In comparison
with the OVA-sensitized mice, the anti-IL-5 treated mice showed
an 80% reduction in upper airway eosinophil infiltration
(P � 0.0005). Mice given IP injections of control rat IgG
developed eosinophil infiltration equal to that of OVA-sensitized/
OVA-challenged mice. (Saline control and intranasal OVA-treated
mice, N � 3 mice per condition. Anti-IL-5 and rat IgG-treated
mice, N � 5 mice per condition).
1056 I. Hussain et al.
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059

demonstration of eosinophils in the ethmoturbinates, but not in
more inferiorly located areas lined by olfactory epithelium,
argues further for an anatomically specific distribution of
eosinophil infiltration. This may reflect differences in the
ability of the respiratory and the olfactory epithelia to mobilize
an allergic inflammatory response.
The epithelium participates in eosinophil recruitment
through production of adhesion molecules and chemokines
[38]. In human allergic rhinitis, an increase in epithelial
expression of ICAM-1 has been observed during natural
allergen exposure [8] and after experimental allergen
challenge [37]. There is little information on ICAM-1
expression in the mouse upper airway, but Asakura et al.
found that topically applied monoclonal anti-ICAM-1 anti-
body caused a significant attenuation of the eosinophil
infiltration into nasal mucosa 24 h after nasal challenge with
OVA in sensitized rats [9]. Furthermore, in a mouse model of
Ova-induced asthma, treatment with anti-ICAM-1 antibody
blocked Ova-inhalation-induced infiltration of eosinophils by
greater than 70% [10]. Similarly, an increase in epithelial
expression of eotaxin has been found in nasal mucosa after
local allergen challenge in patients with allergic rhinitis [11]
and after natural allergen exposure [39]. Intranasal application
of eotaxin has also been shown to increase eosinophils in nasal
secretions in patients with allergic rhinitis [13]. Given these
data, we hypothesized that intranasal OVA challenge may be
associated with an increase in epithelial expression of either
ICAM-1 or eotaxin in OVA-sensitized mice. However, we
were unable to confirm that either of these proteins was
induced in OVA sensitized and challenged mice. Our failure to
detect the induction of either ICAM-1 or eotaxin may reflect a
lack of their induction, insensitivity of the immunostaining
procedure or perhaps the failure to detect an early response.
Certain epitopes of key immunological interest could have
been lost owing to the tissue fixation and/or decalcification
processes employed. Indeed, we found it difficult to detect
lymphocyte surface markers or cytokines in these tissues.
However, this is unlikely to be the case, since we found regions
of constitutive expression of both ICAM-1 and eotaxin in
control mice. Using a similar OVA allergen challenge model,
Rothenberg et al. found that eotaxin mRNA expression
occurred rapidly in the lungs, peaking at 3±6 h [40]. In eotaxin
null mice, the influx of eosinophils into the lungs after allergen
challenge was reduced by 70% at 18 h but was normal 48 h
after allergen challenge [40]. Hence, in our experiments, it is
possible that we could have missed a critical early induction of
epithelial eotaxin expression even though we examined mice
as early as 24 h after allergen challenge. Obviously, more
studies are needed to examine the mechanism of eosinophil
infiltration into the upper airway, including strategies to
selectively inhibit the function of either ICAM-1 or eotaxin in
OVA sensitized and challenged mice.
The development of upper airway eosinophilia required
both systemic OVA sensitization and aerosol or intranasal
OVA challenge. Hence, direct application of OVA to the
upper airway was required to induce eosinophil infiltration.
Intraperitoneal sensitization was found to elicit a greater
increase in bone marrow eosinophils than intranasal
sensitization. This difference in sensitization may explain
the less intense eosinophil infiltration of the upper airway
seen using the intranasal protocol.
Repeated intranasal exposure to OVA has previously been
shown to elicit a weaker sensitization than IP injection with
alum but greater selectivity for eliciting an IgE anti-OVA
response [25]. As a result, the intranasal sensitization protocol
was initially used by Hamelmann et al. to demonstrate IL-5
dependence of lower airway eosinophilia. However, in a
subsequent study, this same group [26] showed that OVA
sensitization by the IP route was also abrogated by anti-IL-5
antibody. Hence, OVA sensitization by either protocol
appears to give rise to a similar IL-5 dependent response.
OVA-induced upper airway eosinophilia was found to be
IL-5-dependent, since administration of IL-5 antibody during
OVA sensitization resulted in an 80% reduction in the
eosinophil response to subsequent OVA challenge. This
finding is consistent with previously published reports. For
example, Okano et al. [41] found that mice sensitized by
repeated intranasal instillation of Schistosoma mansoni egg
antigen (SEA) produced SEA-specific IgE antibody, devel-
oped nasal eosinophilia and had infiltration of the nasal
secretions with lymphocytes that could be demonstrated to
produce IL-5 in vitro. In another study, Asakura et al. [9]
investigated the effect of anti-IL-5 antibody pretreatment on
allergen-induced late-phase nasal eosinophil infiltration in
mice. Consistent with our results, they reported that the nasal
eosinophil response was significantly abrogated by anti-IL-5
mAb pretreatment. In contrast, the numbers of mast cells in
nasal secretions was unaffected by anti-IL-5 mAb. Hence,
similar to mouse models of allergic lower airways disease
[25±27], the mouse upper airway, specifically in areas lined
by respiratory epithelium, is a target for allergic inflamma-
tion, and this process is largely IL-5-dependent.
Most recently, van de Rijn et al. developed a mouse model
of allergic rhinitis in 129/SvEv mice with Aspergillus
fumigatus (Af) antigen [12]. Consistent with our results, they
showed that sensitized mice developed nasal mucosal
eosinophil infiltration after nasal Af challenge. A similar
response was induced in IgE-deficient mice thus suggesting
that IgE is not required for eosinophil infiltration. They also
reported that the upper airway response was characterized by a
paucity of mononuclear cells and a predominance of
eosinophils, whereas comparable studies in the lower airway
have shown a greater admixture of eosinophils and mono-
nuclear cells [12]. Similarly, we detected very few lympho-
cytes (or neutrophils) in the OVA-sensitized, OVA-challenged
nasal tissues. Nonetheless, our results using the anti-IL-5
Upper airway allergic inflammation in mice 1057
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059

antibody (TRFK-5) provide evidence that allergic upper
airway eosinophil infiltration, like that of the lower airway, is
dependent on the elaboration of IL-5. We hope to extend these
observations to a more detailed examination of the participa-
tion of Th2 and Th1 lymphocytes in nasal eosinophil
infiltration using a strategy of passive transfer of OVA-
specific T lymphocytes as recently reported by Randolph and
coworkers [22].
An attractive additional feature of the mouse upper
airway allergen model is the ability to simultaneously
examine mucosal inflammation and bone marrow effects of
allergen sensitization and/or nasal allergen challenge.
Hence, we were able to show that OVA sensitization alone
induced a significant increase in bone marrow eosinophils
but no increase in nasal mucosal eosinophils, and we were
able to show that the IP protocol elicited a stronger
sensitization than the intranasal protocol. By simulta-
neously examining the bone marrow and upper airway
compartments, it may be possible to study in a coordinated
fashion mechanisms of induction of eosinophil bone
marrow progenitor cells and recruitment of these cells into
the upper airway as has been described in the dog [42±44].
To our knowledge, this study represents the first
demonstration of mouse sinus anatomy using MRI images.
Relatively high resolution of sinus mucosal thickness was
achieved with our protocol. In preliminary experiments, we
did not observe any significant change in nasal or sinus
mucosal thickness in response to allergic inflammation,
However, we postulate that MRI imaging may prove useful
in future studies in which more dramatic levels of
inflammation may be seen, such as chronic allergic
inflammation or acute infection. Serial MRI imaging may
offer the advantage of tracking dynamic mucosal changes,
such as the response to treatment, without killing the animal.
Acknowledgements
This work was funded in part by the NIH SCOR grant #
1P50HL54619 (Dr Chaplin) and by the Janssen Pharmaceutica
Research Award in Allergic Rhinitis (Dr Hussain). Dr Chaplin
is an Investigator of the Howard Hughes Medical Institute. The
authors thank Ms. Lisa Fitzgerald for manuscript preparation;
Michael Holtzman for use of Core Laboratory facilities;
Deepak Sampath, Tim Birkland, Brian Faddis, Richard Chole,
Theresa Tolley and Jill Roby for assistance in tissue
preparation and Dwight Look for review of the manuscript.
References
1 Mackay IS, Durham SR. ABC of allergies. Perennial rhinitis.
BMJ 1998; 316 (7135):917±20.
2 Juniper EF. Rhinitis management: the patient's perspective.
Clin Exp Allergy 1998; 28 (Suppl. 6):34±8.
3 Demoly P, Crampette L, Mondain M et al. Assessment of
inflammation in noninfectious chronic maxillary sinusitis. J
Allergy Clin Immunol 1994; 94 (1):95±108.
4 Meltzer EO. The prevalence and medical and economic impact
of allergic rhinitis in the United States. J Allergy Clin
Immunol 1997; 99 (6, part 2):S805±28.
5 Durham SR, Ying S, Varney VA. et al. Cytokine messenger
RNA expression for IL-3, IL-4, IL-5, and granulocyte/
macrophage-colony-stimulating factor in the nasal mucosa
after local allergen provocation: relationship to tissue
eosinophilia. J Immunol 1992; 148 (8):2390±4.
6 Bentley AM, Meng Q, Robinson DS, Hamid Q, Kay AB,
Durham SR. Increases in activated T lymphocytes, eosino-
phils, and cytokine mRNA expression for interleukin-5 and
granulocyte/macrophage colony-stimulating factor in bron-
chial biopsies after allergen inhalation challenge in atopic
asthmatics. Am J Respir Cell Mol Biol 1993; 8 (1):35±42.
7 Robinson DS, Hamid Q, Ying S. et al. Predominant TH2-like
bronchoalveolar T-lymphocyte population in atopic asthma. N
Engl J Med 1992; 326 (5):298±304.
8 Ciprandi G, Pronzato C, Ricca V, Bagnasco M, Canonica GW.
Evidence of intercellular adhesion molecule-1 expression on
nasal epithelial cells in acute rhinoconjunctivitis caused by
pollen exposure. J Allergy Clin Immunol 1994; 94 (4):738±46.
9 Asakura K, Saito H, Watanabe M, Ogasawara H, Matsui T,
Kataura A. Effects of anti-IL-5 monoclonal antibody on the
murine model of nasal allergy. Int Arch Allergy Immunol
1998; 116 (1):49±52.
10 Chin JE, Winterrowd GE, Hatfield CA. et al. Involvement of
intercellular adhesion molecule-1 in the antigen-induced
infiltration of eosinophils and lymphocytes into the airways
in a murine model of pulmonary inflammation. Am J Respir
Cell Mol Biol 1998; 18 (2):158±67.
11 Minshall EM, Cameron L, Lavigne F. et al. Eotaxin mRNA
and protein expression in chronic sinusitis and allergen-
induced nasal responses in seasonal allergic rhinitis. Am J
Respir Cell Mol Biol 1997; 17 (6):683±90.
12 van de Rijn M, Mehlhop PD, Judkins A, Rothenberg ME,
Luster AD, Oettgen HC. A murine model of allergic rhinitis:
studies on the role of IgE in pathogenesis and analysis of the
eosinophil influx elicited by allergen and eotaxin. J Allergy
Clin Immunol 1998; 102 (1):65±74.
13 Hanazawa T, Antuni JD, Kharitonov SA, Barnes PJ. Intranasal
administration of eotaxin increases nasal eosinophils and nitric
oxide in patients with allergic rhinitis. J Allergy Clin Immunol
2000; 105 (1, part 1):58±64.
14 Cowart BJ, Flynn-Rodden K, McGeady SJ, Lowry LD.
Hyposmia in allergic rhinitis. J Allergy Clin Immunol 1993;
91 (3):747±51.
15 Apter AJ, Mott AE, Frank ME, Clive JM. Allergic rhinitis and
olfactory loss. Ann Allergy Asthma Immunol 1995; 75
(4):311±6.
16 Klimek L, Eggers G. Olfactory dysfunction in allergic rhinitis
is related to nasal eosinophilic inflammation. J Allergy Clin
Immunol 1997; 100 (2):158±64.
17 Moll B, Klimek L, Eggers G, Mann W. Comparison of
1058 I. Hussain et al.
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059

olfactory function in patients with seasonal and perennial
allergic rhinitis. Allergy 1998; 53 (3):297±301.
18 Keller A, Margolis FL. Isolation and characterization of rat
olfactory marker protein. J Biol Chem 1976; 251 (20):6232±7.
19 Renz H, Smith HR, Henson JE, Ray BS, Irvin CG, Gelfand
EW. Aerosolized antigen exposure without adjuvant causes
increased IgE production and increased airway responsive-
ness in the mouse. J Allergy Clin Immunol 1992; 89
(6):1127±38.
20 Renz H, Bradley K, Larsen GL, McCall C, Gelfand EW.
Comparison of the allergenicity of ovalbumin and ovalbumin
peptide 323±339. Differential expansion of V beta-expressing
T cell populations. J Immunol 1993; 151 (12):7206±13.
21 Randolph DA, Stephens R, Carruthers CJ, Chaplin DD.
Cooperation between Th1 and Th2 cells in a murine model
of eosinophilic airway inflammation. J Clin Invest 1999; 104
(8):1021±9.
22 Randolph DA, Carruthers CJ, Szabo SJ, Murphy KM, Chaplin
DD. Modulation of airway inflammation by passive transfer of
allergen- specific Th1 and Th2 cells in a mouse model of
asthma. J Immunol 1999; 162 (4):2375±83.
23 Cohn L, Homer RJ, Marinov A, Rankin J, Bottomly K.
Induction of airway mucus production By T helper 2 (Th2)
cells: a critical role for interleukin 4 in cell recruitment but not
mucus production. J Exp Med 1997; 186 (10):1737±47.
24 Kim TS, DeKruyff RH, Rupper R, Maecker HT, Levy S,
Umetsu DT. An ovalbumin-IL-12 fusion protein is more
effective than ovalbumin plus free recombinant IL-12 in
inducing a T helper cell type 1-dominated immune response
and inhibiting antigen-specific IgE production. J Immunol
1997; 158 (9):4137±44.
25 Hamelmann E, Oshiba A, Loader J. et al. Antiinterleukin-5
antibody prevents airway hyperresponsiveness in a murine
model of airway sensitization. Am J Respir Crit Care Med
1997; 155 (3):819±25.
26 Hamelmann E, Cieslewicz G, Schwarze J. et al. Anti-
interleukin 5 but not anti-IgE prevents airway inflammation
and airway hyperresponsiveness. Am J Respir Crit Care Med
1999; 160 (3):934±41.
27 Hamelmann E, Takeda K, Schwarze J, Vella AT, Irvin CG,
Gelfand EW. Development of eosinophilic airway inflamma-
tion and airway hyperresponsiveness requires interleukin-5 but
not immunoglobulin E or B lymphocytes. Am J Respir Cell
Mol Biol 1999; 21 (4):480±9.
28 Collins PD, Marleau S, Griffiths-Johnson DA, Jose PJ,
Williams TJ. Cooperation between interleukin-5 and the
chemokine eotaxin to induce eosinophil accumulation in vivo.
J Exp Med 1995; 182 (4):1169±74.
29 Hardisty JF, Garman RH, Harkema JR, Lomax LG, Morgan
KT. Histopathology of nasal olfactory mucosa from selected
inhalation toxicity studies conducted with volatile chemicals.
Toxicol Pathol 1999; 27 (6):618±27.
30 Ishida M, Amesara R, Ukai K, Sakakura Y. Antigen (DNP-As)
-induced allergic rhinitis model in guinea pigs. Ann Allergy
1994; 72 (3):240±4.
31 Ressler KJ, Sullivan SL, Buck LB. A zonal organization of
odorant receptor gene expression in the olfactory epithelium.
Cell 1993; 73 (3):597±609.
32 Blatt EN, Yan XH, Wuerffel MK, Hamilos DL, Brody SL.
Forkhead transcription factor HFH-4 expression is temporally
related to ciliogenesis. Am J Respir Cell Mol Biol 1999; 21
(2):168±76.
33 Keller A, Margolis FL. Immunological studies of the rat
olfactory marker protein. J Neurochem 1975; 24 (6):1101±6.
34 Mlynek ML. Comparative investigations on the specificity of
Adams' reaction and the Biebrich scarlet stain for the
demonstration of eosinophilic granules. Klin Wochenschr
1985; 63 (14):646±7.
35 Schaefer HE, Fischer R. Specific staining of eosinophilic
granulocytes with Biebrich scarlet. Klin Wochenschr 1968; 46
(7):396±7.
36 Daly LE, Bourke GJ, McGilvray JW. Interpretation and uses
of medical statistics. 4th edn. Oxford. Boston. Blackwell
Scientific Publications., 1991.
37 Ciprandi G, Ricca V, Passalacqua G, Fasolo A, Canonica GW.
Intranasal fluticasone propionate reduces ICAM-1 on nasal
epithelial cells both during early and late phase after allergen
challenge. Clin Exp Allergy 1998; 28 (3):293±9.
38 Rothenberg ME, Eosinophilia. N Engl J Med 1998; 338
(22):1592±600.
39 Pullerits T, Linden A, Praks L, Cardell LO, Lotvall J.
Upregulation of nasal mucosal eotaxin in patients with allergic
rhinitis during grass pollen season: effect of a local
glucocorticoid. Clin Exp Allergy 2000; 30 (10):1469±75.
40 Rothenberg ME, MacLean JA, Pearlman E, Luster AD, Leder
P. Targeted disruption of the chemokine eotaxin partially
reduces antigen-induced tissue eosinophilia. J Exp Med 1997;
185 (4):785±90.
41 Okano M, Nishizaki K, Abe M. et al. Strain-dependent
induction of allergic rhinitis without adjuvant in mice. Allergy
1999; 54 (6):593±601.
42 Inman MD, Denburg JA, Ellis R, Dahlback M, O'Byrne PM.
Allergen-induced increase in bone marrow progenitors in
airway hyperresponsive dogs: regulation by a serum hemopoie-
tic factor. Am J Respir Cell Mol Biol 1996; 15 (3):305±11.
43 Inman MD, Ellis R, Wattie J, Denburg JA, O'Byrne PM.
Allergen-induced increase in airway responsiveness, airway
eosinophilia, and bone-marrow eosinophil progenitors in mice.
Am J Respir Cell Mol Biol 1999; 21 (4):473±9.
44 Woolley MJ, Denburg JA, Ellis R, Dahlback M, O'Byrne PM.
Allergen-induced changes in bone marrow progenitors and
airway responsiveness in dogs and the effect of inhaled
budesonide on these parameters. Am J Respir Cell Mol Biol
1994; 11 (5):600±6.
45 Mery S, Gross EA, Joyner DR, Godo M, Morgan KT. Nasal
diagrams: a tool for recording the distribution of nasal lesions
in rats and mice. Toxicol Pathol 1994; 22 (4):353±72.
Upper airway allergic inflammation in mice 1059
q 2001 Blackwell Science Ltd, Clinical and Experimental Allergy, 31, 1048±1059
Related Documents


![Research Paper Deguelin Attenuates Allergic Airway ... · Research Paper Deguelin Attenuates Allergic Airway Inflammation via ... pathophysiology of asthma [4]. ... schematic diagram](https://static.cupdf.com/doc/110x72/5b396a827f8b9ab9068e82d4/research-paper-deguelin-attenuates-allergic-airway-research-paper-deguelin.jpg)








