Lecture 1 & 2 January 2011 1 2011 January 4 & 6 MIMM314 R G E Palfree IMMUNOLOGY Year 2011 version Dr. Roger Palfree’s Lectures 1 & 2 Lecture 1 begins with Introduction to the course (not included here) Lectures 1cntd and 2 Introduction to Immunology The primary course textbook was Janeway’s Immunobiology (Garland Science). Get it, read it, and you will likely keep it for reference for many years. Made available via www.rogerpalfree.com 2011 January 4 & 6 MIMM314 R G E Palfree IMMUNOLOGY Lecture 1 Part B Introduction to Immunology Challenges for a Living System Defence Strategies and Mechanisms Immunity and Immunology Immune System Overview

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Lecture 1 & 2 January 2011
1
2011 January 4 & 6 MIMM314 R G E Palfree
IMMUNOLOGY
Year 2011 version
Dr. Roger Palfree’s Lectures 1 & 2
Lecture 1 begins withIntroduction to the course (not included here)
Lectures 1cntd and 2Introduction to Immunology
The primary course textbook was Janeway’s Immunobiology (Garland Science).Get it, read it, and you will likely keep it for reference for many years.
Made available via www.rogerpalfree.com
2011 January 4 & 6 MIMM314 R G E Palfree
IMMUNOLOGY
Lecture 1 Part B
Introduction to Immunology
� Challenges for a Living System
� Defence Strategies and Mechanisms
� Immunity and Immunology
� Immune System Overview

Lecture 1 & 2 January 2011
2
Rx
2011 January 4 & 6 MIMM314 R G E Palfree
Challenges for a Living System
usedby
system
waste
activeuptake
activeejection
Rx
Rx
mustreproduce
MaintenanceGrowthMovementAdaptationReproduction
2011 January 4 & 6 MIMM314 R G E Palfree
Challenges for a Living System
Generally the same for organisms which are• single cells or multi-cellular• actively mobile or passively mobile
(Note that plants have mobile stages in the scattering of seeds or even the vegetative reproduction such as via rhizomes or runners)
Some survival strategies are commonOthers vary widely from one system to another
Species survival does not require the survival of all individuals

Lecture 1 & 2 January 2011
3
2011 January 4 & 6 MIMM314 R G E Palfree
Survival Strategies against other Living Systems
Against Competitors• Chase them away• Kill them• Even better, eat them
Against Predators• Run• Hide• Make it not worth the effort (e.g. fight, be unpalatable) • Be reproductively prolific
Against Microbes• Keep them out• Poison them• Use them as food• Die to reduce their ability to proliferate and spread
Prisoner Zerofrom
Dr. Who
2011 January 4 & 6 MIMM314 R G E Palfree
Defence Strategies against Microbes
Barrier• Prevent entry• Prevent adherence• Wash them away• Antimicrobial chemicals
Non-Self RecognitionSoluble receptors• Hamper adhesion• Control uptake and clearance• Signalling for coordinated defencesSurface receptors• Control uptake and clearance• Signalling for coordinated defences
Production of Toxic ChemicalsCompartments for inactivation and breakdownControlled Death

Lecture 1 & 2 January 2011
4
2011 January 4 & 6 MIMM314 R G E Palfree
Innate Defences, Pathogens and Immunity
Innate Defence is provided by mechanisms and receptors for common non-self (or altered self), which have evolved and have been selected because they have improved survival of the species.
Pathogens have evolved to evade some innate defence mechanisms.
Immunity was recognized a long time before disease was understood.
• People who had survived a contagious disease were not affected by it during a subsequent epidemic.
The scientific study of this phenomenon became Immunology.
Immunological understanding of Immunity: Recruitment of defence mechanisms with novel, pathogen-specific receptors.
2011 January 4 & 6 MIMM314 R G E Palfree
Innate Defence and the Immune System
In the rest of this lecture we are concerned primarily with the mammalian immune system (which includes innate defence system and the adaptive response system which provides individual immunity).
We will :
• Look at some components of the immune system• organs, cells, molecules
• Look a bit more closely at some of the functions of immune subsystems
• Touch on some of the work of early pioneers in immunology
• Have an overview and quick tour through some of the material to be covered in detail later in the course.

Lecture 1 & 2 January 2011
5
2011 January 4 & 6 MIMM314 R G E Palfree
The Skin
Skin:
accounts for about 16% of adult total body weight
From: http://www.mederma.com/ce3.html
Besides providing a
continually renewing barrier
to microorganisms, skin also
actively challenges microbes
through sweat components.
Antimicrobial peptides are
produced by skin cells and
secreted, esp. upon damage.
Lipid is secreted and
transformed into a
hydrophobic barrier called the
stratum corneum.
2011 January 4 & 6 MIMM314 R G E Palfree
Mucosal Surfaces
Mucosal surfaces - a vast surface area
Fluid flow carries microbes away from vulnerable surfaces
Non-specific surfactants and specific innate (SP-A, SP-D)
and adaptive (IgA) receptors in the mucus reduce microbe adhesion
to the epithelial cells, opsonize for phagocytosis (e.g. by alveolar
macrophage in lung), and neutralize toxins (esp. IgA).
Enzymes and antimicrobial peptides attack microbes and
toxins. Many microbes will not survive the low pH in the stomach
The tissue just beneath the epithelial layer contains
structures which promote efficient antigen presentation to
lymphocytes. Broadly called
the MucosaMucosa--Associated Lymphoid Tissue Associated Lymphoid Tissue -- MALTMALT

Lecture 1 & 2 January 2011
6
2011 January 4 & 6 MIMM314 R G E Palfree
Scenario: Skin wound deep enough to damage blood vessels
Platelets adhere to collagen outside the blood vessel. They become activated and release the contents of stored granules
activate other platelets.
Activated platelets bind to fibrinogen, which leads to their aggregation.
The clotting cascade is initiated when clotting factor VII comes into contact with tissue factor on cell surfaces outside the blood vessel.
Activated platelets and other cells provide a phospholipid surface for some clotting reactions.
Thrombin converts fibrinogen to fibrin and the fibrin becomes cross-linked into a meshwork in which aggregated platelets are trapped.
Activated platelets and the clotting cascade produce inflammatory mediators which act on blood vessels and leukocytes.
2011 January 4 & 6 MIMM314 R G E Palfree
Inflammation (local heat, edema, white blood cells)
Inflammatory mediators are produced
� in the clotting process� by activated platelets� by cells responding to tissue damage� by cells responding to microbial material entering the wound� in the complement cascade activated by microbes or immune
complexes
(We will look at the complement system in detail later in the course.)
Inflammation includes recruitment of leukocytes to the wound.
These cells are involved in removal of dammaged tissue and killing of microorganisms.
They also provide signals which promote further inflammation in the case of infection, or promote wound healing.

Lecture 1 & 2 January 2011
7
2011 January 4 & 6 MIMM314 R G E Palfree
Innate immunity protects immediately, or generates a maximal
response very quickly (minutes to several hours).
Adaptive immunity requires several days to deliver high-
affinity antibodies or antigen-specific effector T cells to the
sites of infection.
A bacterium doubling every 20 minutes would produce 1045
progeny in 50 hours. The innate immune system must at least
slow it down to an effective doubling time of several hours, or
the host would die before adaptive immunity could develop !
By the way, 1045 E. coli bacteria would weigh 1030 kg
So an infected overweight , 100 kg human would not provide sufficient nutrient for such a sustained growth rate even in the absence of innate
defence mechanisms.
2011 January 4 & 6 MIMM314 R G E Palfree
If the wound becomes infected, the adaptive immune system gets involved:
There is enough microbial material to activate dendritic cells.
�Dendritic cells are in the tissues�Monocytes recruited from the circulation mostly differentiate to macrophage�Some monocytes differentiate into dendritic cells
Dendritic cells take up and process foreign material to present to T lymphocytes.
Activated dendritic cells migrate to the nearby lymph nodes
T lymphocytes circulate from the blood into lymph nodes, through lymphatic vessels, and back into the blood.
Any T lymphocyte with a receptor which binds a foreign peptide presented by an activated dendritic cell will become activated, proliferate, and differentiate into effector T cells.

Lecture 1 & 2 January 2011
8
2011 January 4 & 6 MIMM314 R G E Palfree
Effector T cells
If they are T helper type 2 cells, they will interact with B lymphocytes which pass through the lymph node.
They help activate any B cell which has a receptor for foreign material of the type which was the source of the peptide which triggered the T cell activation.
Activated B cells proliferate and differentiate to produce antibodies.
Other effector T cells enter the blood, circulate, and are recruited by inflammatory mediators and cytokines to the site of infection.
T helper type 1 cells interact with macrophage to make them kill and clear pathogen.
Cytotoxic T cells kill cells which have become infected with intracellular pathogens.
2011 January 4 & 6 MIMM314 R G E Palfree
Before we look at these cells, tissues and processes in more detail and develop concepts to carry into future lectures, I would like to step back to early days and look at some of the key people, discoveries , innovations and theories which brought immunology to maturity.

Lecture 1 & 2 January 2011
9
2011 January 4 & 6 MIMM314 R G E Palfree
Development and Application of Scientific Method and
Theory to the Study and Manipulation of Immunity
The following slides point to some of the key contributors in the development of immunology as a science.
The contributions noted here do not constitute an exhaustive list.
( You can expect an exam question from this historical review )
2011 January 4 & 6 MIMM314 R G E Palfree
http://www.nursingceu.com/NCEU/courses/smallpox/
Smallpox
Development of smallpox in a child who survived.
Photos taken 5, 7, 10, and 15 days after the appearance of a rash.
Blisters will turn into highly contagious scabs, which fall off the body, leaving the victim permanently scarred and in some cases blind.
Lecture 1 & 2 January 2011
10
2011 January 4 & 6 MIMM314 R G E Palfree
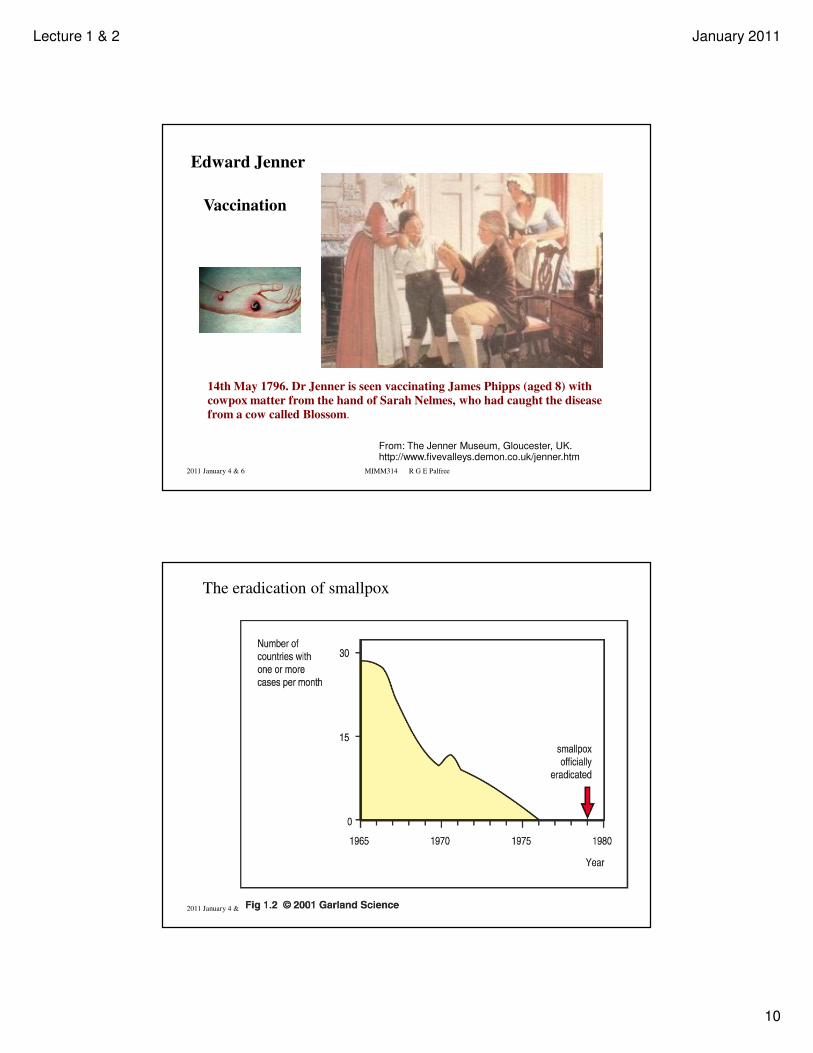
14th May 1796. Dr Jenner is seen vaccinating James Phipps (aged 8) with
cowpox matter from the hand of Sarah Nelmes, who had caught the disease
from a cow called Blossom.
From: The Jenner Museum, Gloucester, UK. http://www.fivevalleys.demon.co.uk/jenner.htm
Vaccination
Edward Jenner
2011 January 4 & 6 MIMM314 R G E Palfree
The eradication of smallpox

Lecture 1 & 2 January 2011
11
2011 January 4 & 6 MIMM314 R G E Palfree
Spallanzani
Supporting the belief in spontaneous generation of living things, John Needham had heated various organic substances in a flask hermetically closed to sterilize them. After treatment, microbes emerged in all the solutions.
In 1768 Lazzaro Spallanzani repeated this experiment but used a higher temperature and longer sterilization time. No microbial growth occurred. Also microbe-containing media were sterilized by his procedure.
Despite this demonstration, the popular belief in spontaneous generation was not quickly abandoned, and even supported by some eminent scientists up to the 1860s
2011 January 4 & 6 MIMM314 R G E Palfree
Robert Koch (1843-1910)
http://www.nobel.se/medicine/laureates/1905/koch-bio.html
Brilliant investigator.
Stains for microbes.
Developed methods like use of solid nutrient media for microbial growth and cloning.
Showed that microbes cause diseases.
Identified several microbial pathogens.

Lecture 1 & 2 January 2011
12
2011 January 4 & 6 MIMM314 R G E Palfree
Louis Pasteur (1822-1895)
http://www.physics.ucla.edu/class/85HC_Gruner/bios/pasteur.html
Preeminent scientist esp: microbiologist.
Work on fermentation. Contributed to demise of belief in spontaneous generation, and to the proof of microbes as agents of disease.
Produced vaccines against cholera in chickens, and against rabies.
2011 January 4 & 6 MIMM314 R G E Palfree
Elie Metchnikoff (1845-1916)
From http://www.pasteur.fr/infosci/archives/mtc0.html
Discovered phagocytosis while studying starfish larvae.
It was a common belief that the leukocytes in wounds were bad, being carriers of microbes into the body.
He argued that the so-called macro- and microphages were important effectors of immunity.
(He considered antibodies to be helpers of phagcytosis.)
Lecture 1 & 2 January 2011
13
2011 January 4 & 6 MIMM314 R G E Palfree
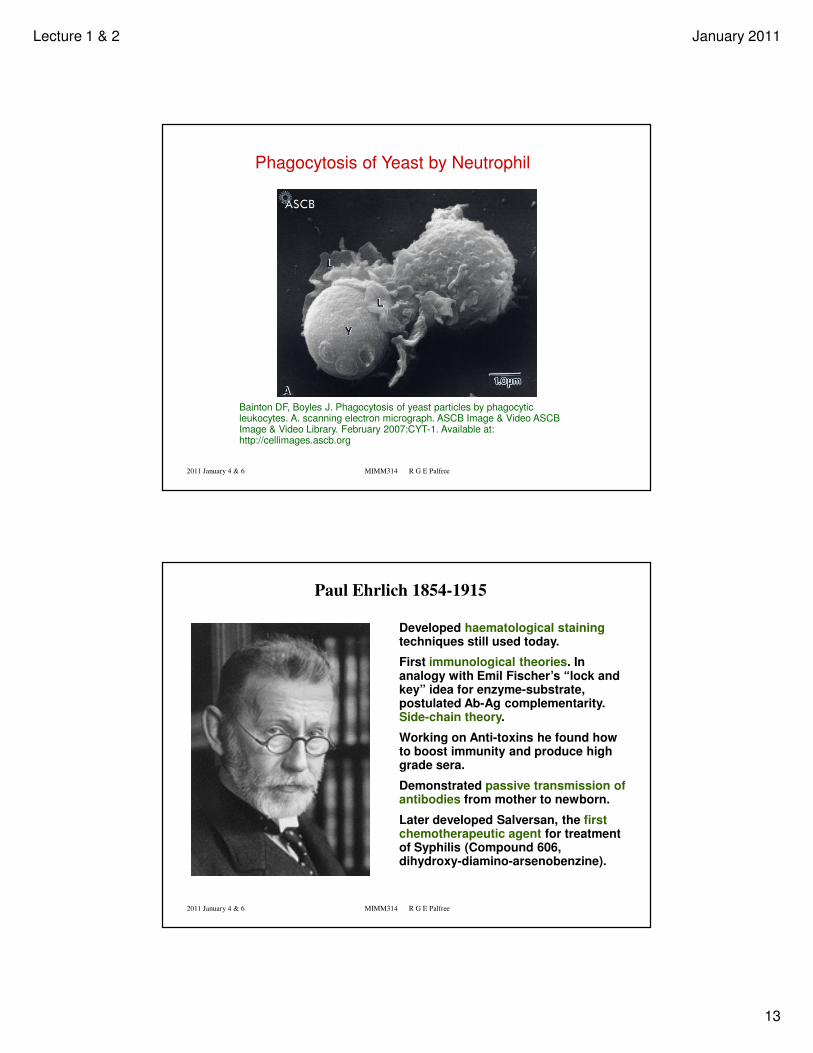
Phagocytosis of Yeast by Neutrophil
Bainton DF, Boyles J. Phagocytosis of yeast particles by phagocytic leukocytes. A. scanning electron micrograph. ASCB Image & Video ASCB Image & Video Library. February 2007:CYT-1. Available at: http://cellimages.ascb.org
2011 January 4 & 6 MIMM314 R G E Palfree
Paul Ehrlich 1854-1915
Developed haematological stainingtechniques still used today.
First immunological theories. In analogy with Emil Fischer’s “lock and key” idea for enzyme-substrate, postulated Ab-Ag complementarity. Side-chain theory.
Working on Anti-toxins he found how to boost immunity and produce high grade sera.
Demonstrated passive transmission of antibodies from mother to newborn.
Later developed Salversan, the first chemotherapeutic agent for treatment of Syphilis (Compound 606, dihydroxy-diamino-arsenobenzine).

Lecture 1 & 2 January 2011
14
2011 January 4 & 6 MIMM314 R G E Palfree
Serum Proteins
IgG a γ-globulin
2011 January 4 & 6 MIMM314 R G E Palfree
Behring together with his colleagues Wernicke (left) and Frosch (center) in Robert Koch's laboratory in Berlin.
Emil von Behring (1854-1917): The Founder of Serum Therapy(injection of serum from immune animal to cure disease in another)
Photo: Aventis Behring From: http://www.nobel.se/medicine/articles/behring/

Lecture 1 & 2 January 2011
15
2011 January 4 & 6 MIMM314 R G E Palfree
Shibasaburo Kitasato(1892-1931)
Photo: from Aventis Behring
Worked with Behring in developing an effective therapeutic serum against tetanus for passive immunization:
The particularly poisonous substances from bacteria – or toxins -could be rendered harmless by the serum of animals immunized with attenuated forms of the infectious agent through “antidotes” or “antitoxins”.
2011 January 4 & 6 MIMM314 R G E Palfree
Historical engraving showing how the medicinal serum
was obtained from immunized horses.
Photo: from Aventis Behring
Lecture 1 & 2 January 2011
16
2011 January 4 & 6 MIMM314 R G E Palfree
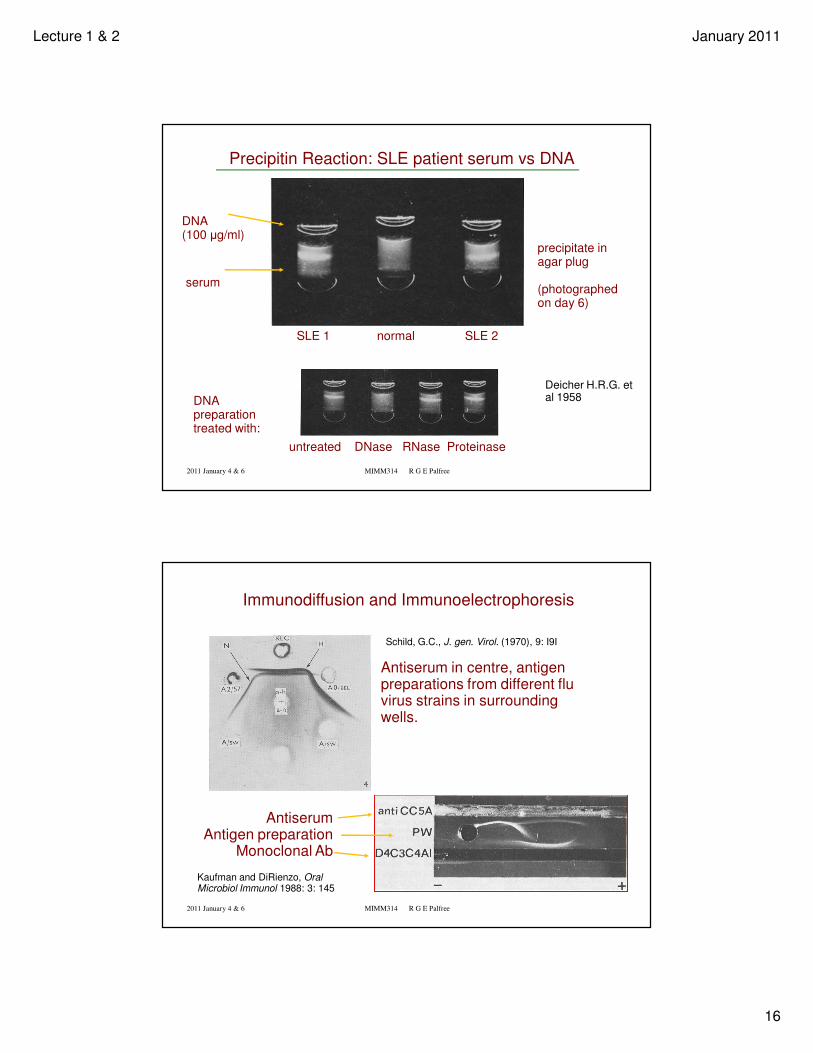
Precipitin Reaction: SLE patient serum vs DNA
DNA (100 µg/ml)
serum
SLE 1 normal SLE 2
precipitate in agar plug
(photographed on day 6)
DNA preparation treated with:
untreated DNase RNase Proteinase
Deicher H.R.G. et al 1958
2011 January 4 & 6 MIMM314 R G E Palfree
Schild, G.C., J. gen. Virol. (1970), 9: I9I
Immunodiffusion and Immunoelectrophoresis
Kaufman and DiRienzo, Oral Microbiol Immunol 1988: 3: 145
Antiserum in centre, antigen preparations from different flu virus strains in surrounding wells.
AntiserumAntigen preparation
Monoclonal Ab

Lecture 1 & 2 January 2011
17
2011 January 4 & 6 MIMM314 R G E Palfree
http://www.bact.wisc.edu/Microtextbook/index.php?module=Book&func=displayarticle&art_id=270
Immuno-agglutinationSerological reactions for Salmonella isolates(-) The negative reaction to an anti-B serum.
(+) The positive reaction is to an anti-C1 serum. Note the grainy appearance of the positive reaction.
These are just bubbles
Agglutinated bacteria
2011 January 4 & 6 MIMM314 R G E Palfree
In 1899 showed that serum from immunized animals contained not only molecules (antibodies) which specifically bind to the foreign substance causing agglutination or precipitation, but also a heat-labile substance found in normal blood, which gets fixed to the immune complex and is necessary for killing some microbes.
He called the heat labile substance Alexine, but it is now called Complement – it complements the activity of antibodies.
Jules Bordet (1870-1961)
From: http://www.pasteur.be/bordetuk.htm

Lecture 1 & 2 January 2011
18
2011 January 4 & 6 MIMM314 R G E Palfree
What complement can do to a red blood cell
We will deal with complement in detail later.
2011 January 4 & 6 MIMM314 R G E Palfree
Ray Owen
In 1940s - Showed that genetically different twin calves, which had shared blood circulation during development in utero, were immunologically tolerantof tissues grafted from the other twin.
This required attention to mechanisms
whereby the immune system
distinguished self from non-self.
Photograph taken from Caltech’s oral histories online:
http://oralhistories.library.caltech.edu/123/01/Owen_OHO.pdf

Lecture 1 & 2 January 2011
19
2011 January 4 & 6 MIMM314 R G E Palfree
Microbiology Department, University of Melbourne, 1965
Sir Macfarlane Burnet
From: http://www.wehi.edu.au/collegiate/ASI/burnet.html
Clonal Selection:
In 1950s postulated the preexistence of many different potential antibody-producing cells which display surface membrane-bound forms of the antibody it could be activated to produce.
Also proposed clonal deletion (See later slide)
2011 January 4 & 6 MIMM314 R G E Palfree
During the 1950s, James Gowans revealed the life cycle of the lymphocyte, whose role was at that time completely obscure. Gowans showed that the small lymphocyte continuously recirculated from the blood to the lymph and back again to the blood. He later demonstrated that this cell was at the centre of the (adaptive) immunological response.
James Gowans
From: http://www.path.ox.ac.uk/history.htm
Lecture 1 & 2 January 2011
20
2011 January 4 & 6 MIMM314 R G E Palfree
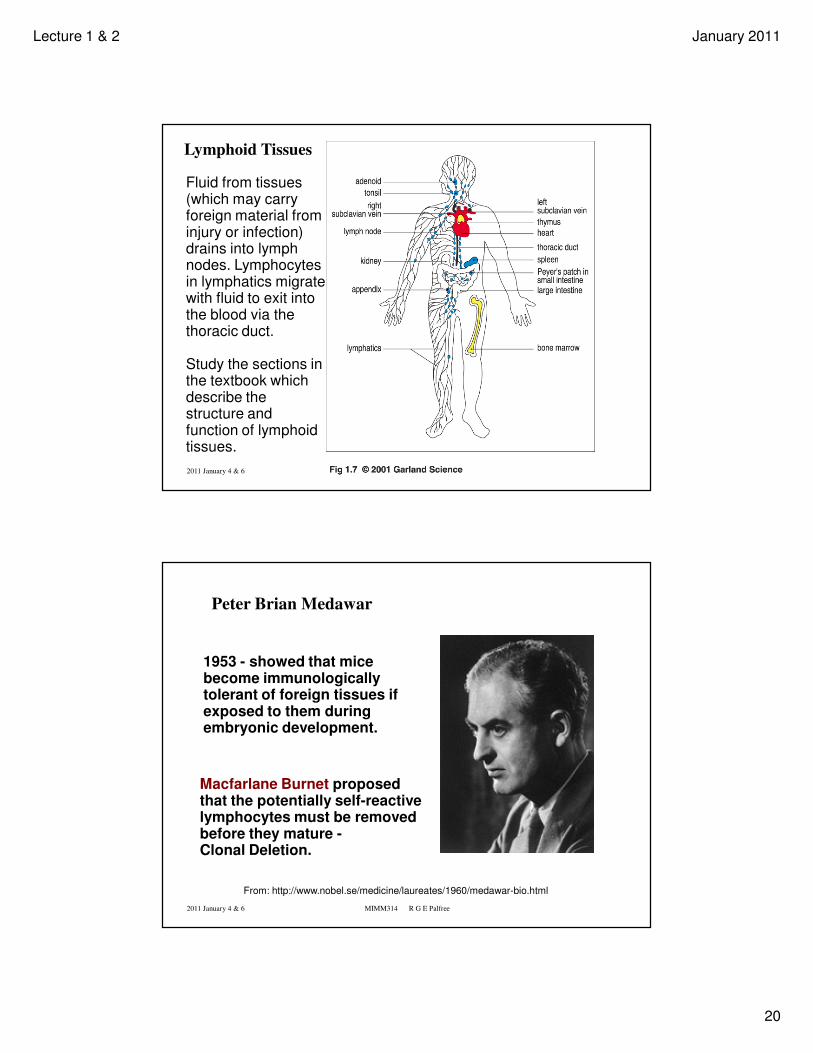
Lymphoid Tissues
Fluid from tissues (which may carry foreign material from injury or infection) drains into lymph nodes. Lymphocytes in lymphatics migrate with fluid to exit into the blood via the thoracic duct.
Study the sections in the textbook which describe the structure and function of lymphoid tissues.
2011 January 4 & 6 MIMM314 R G E Palfree
From: http://www.nobel.se/medicine/laureates/1960/medawar-bio.html
Peter Brian Medawar
1953 - showed that mice become immunologically tolerant of foreign tissues if exposed to them during embryonic development.
Macfarlane Burnet proposed that the potentially self-reactive lymphocytes must be removed before they mature -Clonal Deletion.

Lecture 1 & 2 January 2011
21
2011 January 4 & 6 MIMM314 R G E Palfree
George D. Snell
1929: Clarence Little founded the Jackson Memorial Laboratory, Bar Harbor, Maine, to study transplantable tumors in genetically homogeneous animals.
1935: George Snell was recruited to the Jackson Laboratory. By meticulous inbreeding and testing of mice to obtain strains differing only at a single genetic locus, he “invented” the congenic mouse strain.
2011 January 4 & 6 MIMM314 R G E Palfree
Proceedings of the American Philosophical Society, vol. 145
(3), September 2001
Through this work Snelldiscovered a genetic locus strongly connected with graft rejection. He labelled it “H” for histocompatibility.
Peter Gorer discovered antibody involved in rejection of a tumor graft, and called the antigen “antigen II”. It mapped to the H locus, so the locus became known as H-2.

Lecture 1 & 2 January 2011
22
2011 January 4 & 6 MIMM314 R G E Palfree
Peter Gorer (1907-1961), at Guy’s Hospital, London, working on mice, discovered antibody which was involved in rejection of a tumor graft, and called the target antigen “antigen II”. It mapped to the H locus, so the locus became known as H-2.
We now know it is more complicated than a single
gene, and we refer to the set of genes as the Major
Histocompatibility Complex (MHC).
But in mice, the H-2 label remains. Thus, for
example, H2K and H2D are mouse class I MHC
molecules encoded by genes within the H-2 locus.
2011 January 4 & 6 MIMM314 R G E Palfree
WHO IS THE FATHER OF IMMUNOLOGY ?
• “Edward Jenner is acknowledged as the Father of Immunology”
• “Paul Ehrlich has been regarded as a founding father of immunology”
• “Sir Frank Macfarlane Burnet ‘The Father of Immunology’”
• “Louis Pasteur: ‘Father of immunology and microbiology.’”
• “The ‘father’ of immunology , Dr. Emil Adolph von Behring”
• “Elias Metchnikoff ... can justly be considered the father of immunology”
• “Dr. George Snell (1903-1996): scientists ... called him the father of immunology.”.

Lecture 1 & 2 January 2011
23
2011 January 4 & 6 MIMM314 R G E Palfree
Some of the slides in this presentation were taken from“Cellular and Molecular Immunology”, Abbas, Lichtman, Pober, 4th Edn., W.B.
Saunders Co. "Immunobiology: The immune system in health and disease”, Janeway, Travers,
Walport, Shlomchik, 2001, 5th Edition, Garland Publishing Inc.
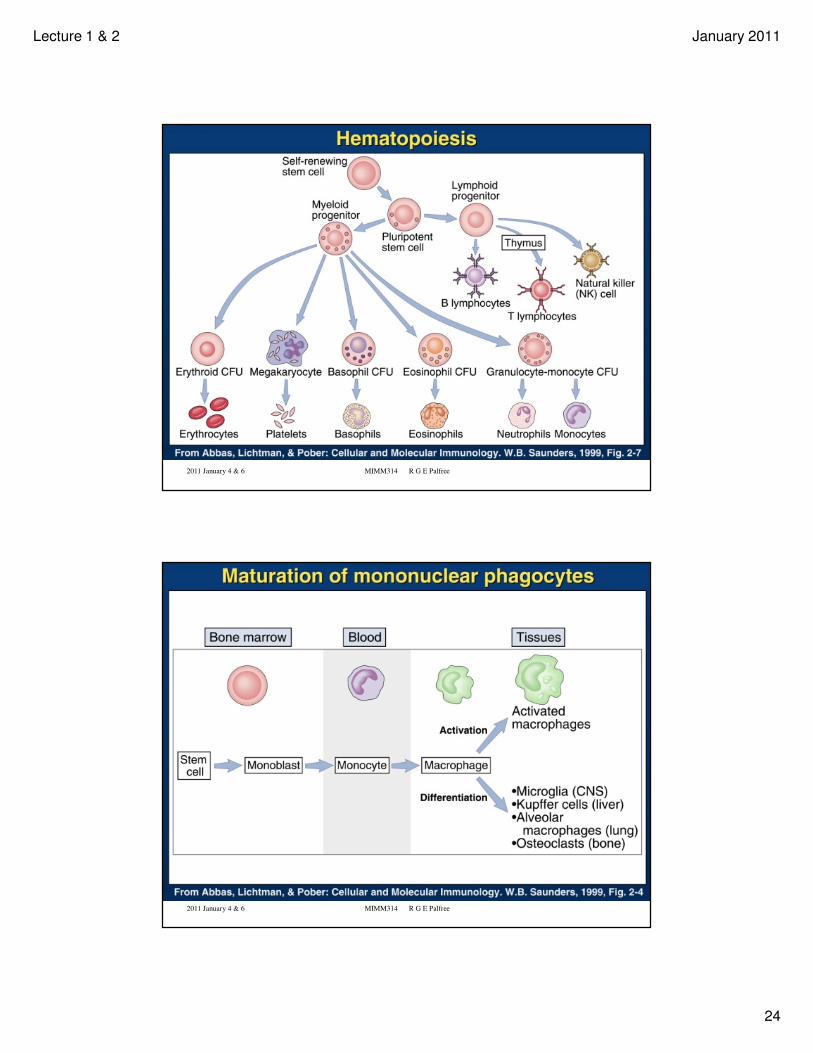
• Schematic of hematopoietic lineages
• Effector functions of the cell types
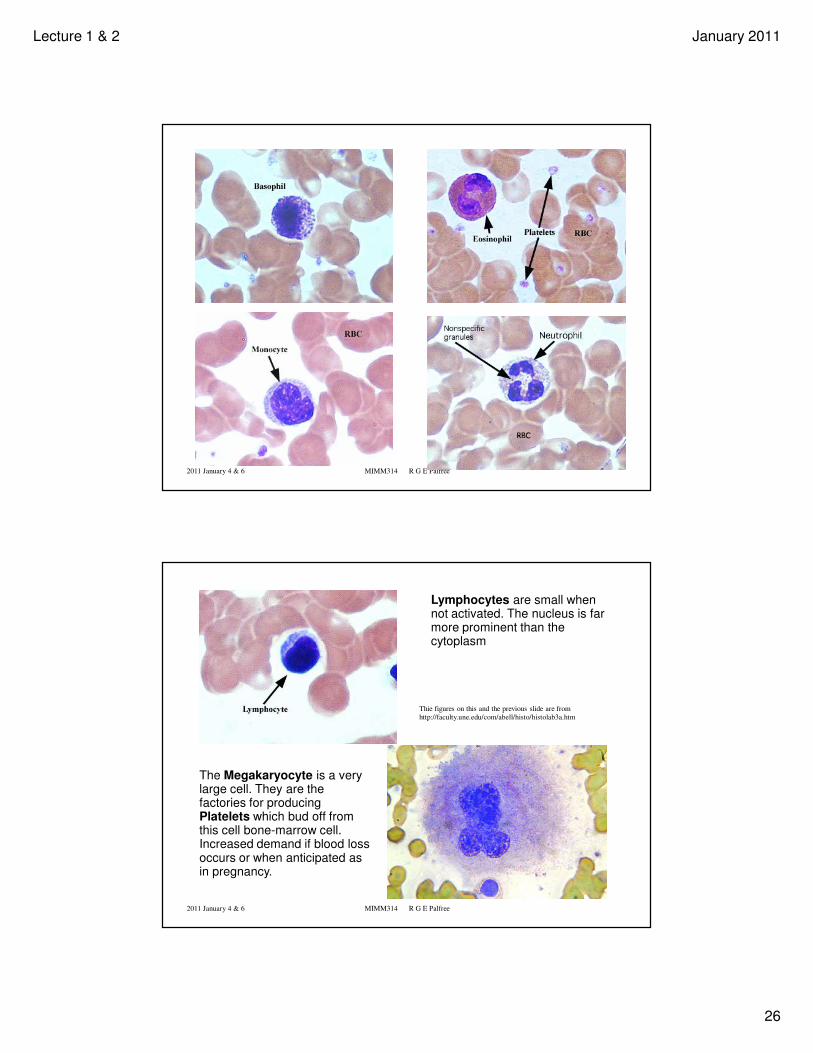
• Some prettier pictures showing characteristic appearance of stained basophil,
eosinophil, monocyte, neutrophil, lymphocite, megakaryocyte. (These were taken
from The University of New England, College of Osteopathic Medicine: Anatomy 503,
Human Histology, by Allen L. Bell http://faculty.une.edu/com/abell/histo/histolab2.htm ).
• Types of effector T lymphocyte
Hematopoietic Cells and the Immune System -
Overview and Refresher
2011 January 4 & 6 MIMM314 R G E Palfree
Overview and Preview
The following slides go quickly through an introduction to hematopoietic cells, which you should read about in Ch. 1 of the Immunobiology text.
Then they draw from later in the course to give you an immediate introduction to some basic mechanisms of adaptive immunity*.
* This provides functional context for our discussion of lymphoid tissues,
and allows better understanding of concepts concerning the integration of the innate and adaptive systems. Exam questions on details of antigen processing & presentation, T cell & B cell development & activation, and gene rearrangement for generating the antigen-receptor repertoir will come from the later lectures.
Lecture 1 & 2 January 2011
24
2011 January 4 & 6 MIMM314 R G E Palfree
2011 January 4 & 6 MIMM314 R G E Palfree
Lecture 1 & 2 January 2011
25
2011 January 4 & 6 MIMM314 R G E Palfree
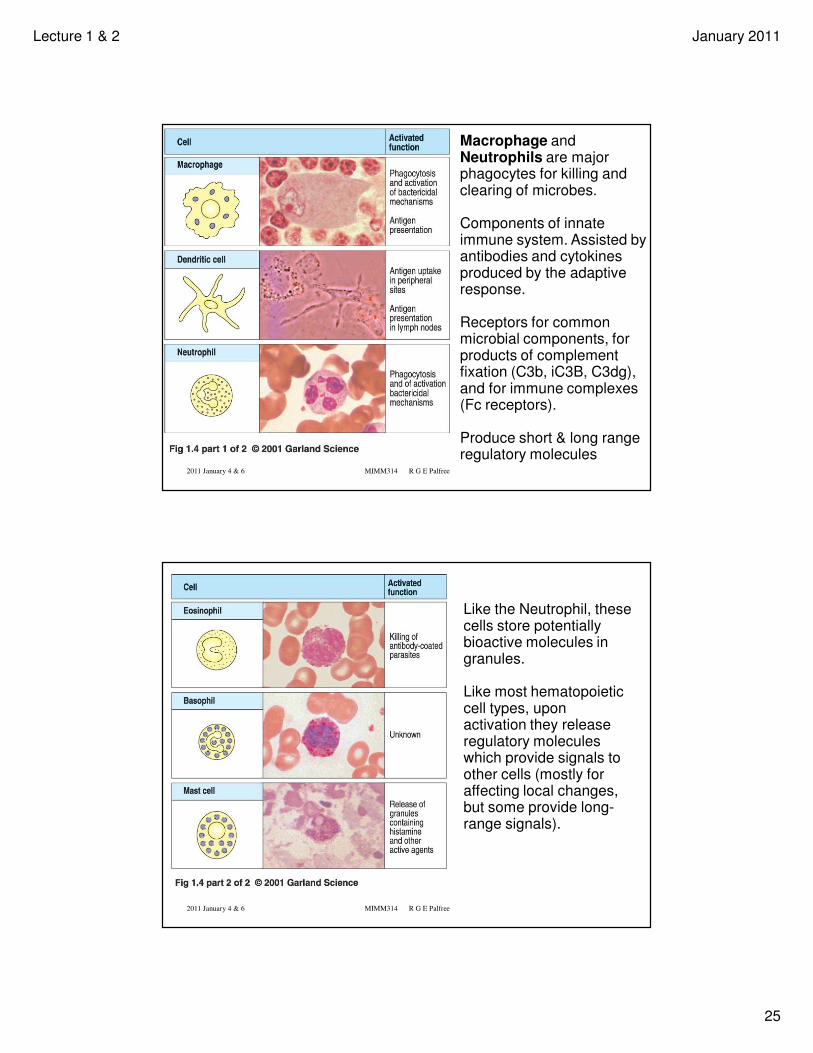
Macrophage and Neutrophils are major phagocytes for killing and clearing of microbes.
Components of innate immune system. Assisted by antibodies and cytokines produced by the adaptive response.
Receptors for common microbial components, for products of complement fixation (C3b, iC3B, C3dg), and for immune complexes (Fc receptors).
Produce short & long range regulatory molecules
2011 January 4 & 6 MIMM314 R G E Palfree
Like the Neutrophil, these cells store potentially bioactive molecules in granules.
Like most hematopoietic cell types, upon activation they release regulatory molecules which provide signals to other cells (mostly for affecting local changes, but some provide long-range signals).
Lecture 1 & 2 January 2011
26
2011 January 4 & 6 MIMM314 R G E Palfree
2011 January 4 & 6 MIMM314 R G E Palfree
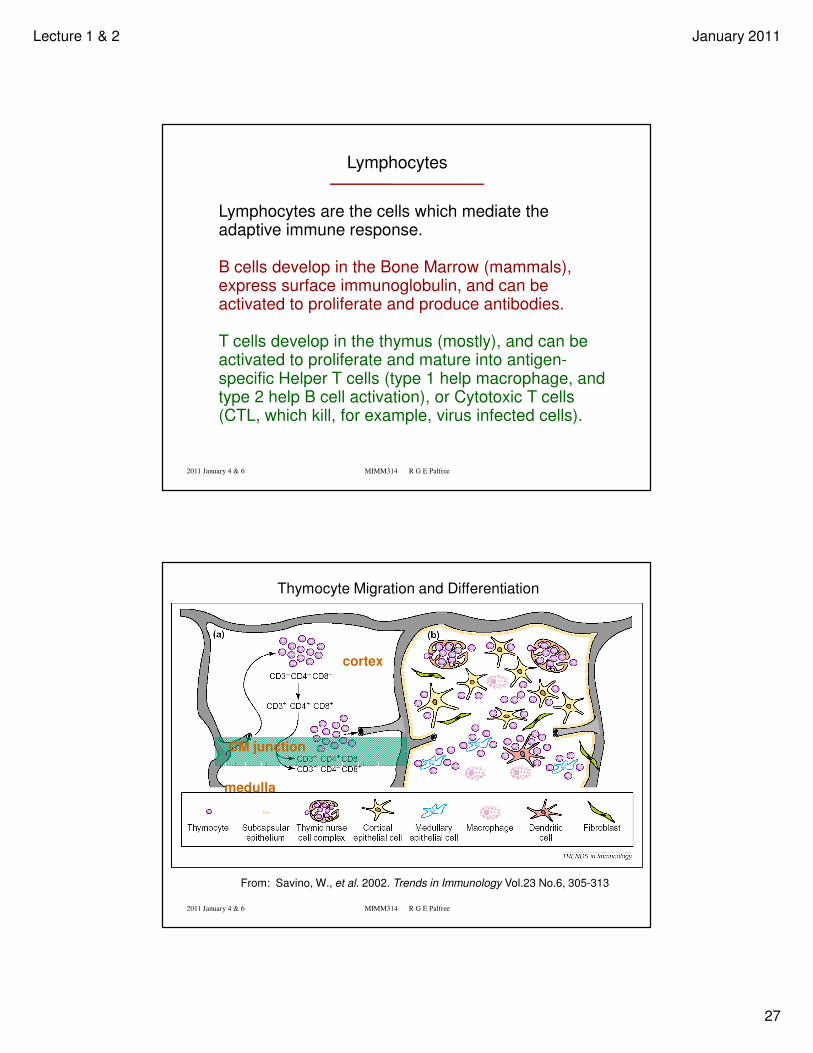
Lymphocytes are small when not activated. The nucleus is far more prominent than the cytoplasm
The Megakaryocyte is a very large cell. They are the factories for producing Platelets which bud off from this cell bone-marrow cell. Increased demand if blood loss occurs or when anticipated as in pregnancy.
Thie figures on this and the previous slide are from
http://faculty.une.edu/com/abell/histo/histolab3a.htm
Lecture 1 & 2 January 2011
27
2011 January 4 & 6 MIMM314 R G E Palfree
Lymphocytes
Lymphocytes are the cells which mediate the adaptive immune response.
B cells develop in the Bone Marrow (mammals), express surface immunoglobulin, and can be activated to proliferate and produce antibodies.
T cells develop in the thymus (mostly), and can be activated to proliferate and mature into antigen-specific Helper T cells (type 1 help macrophage, and type 2 help B cell activation), or Cytotoxic T cells (CTL, which kill, for example, virus infected cells).
2011 January 4 & 6 MIMM314 R G E Palfree
From: Savino, W., et al. 2002. Trends in Immunology Vol.23 No.6, 305-313
Thymocyte Migration and Differentiation
cortex
medulla
CM junction

Lecture 1 & 2 January 2011
28
2011 January 4 & 6 MIMM314 R G E Palfree
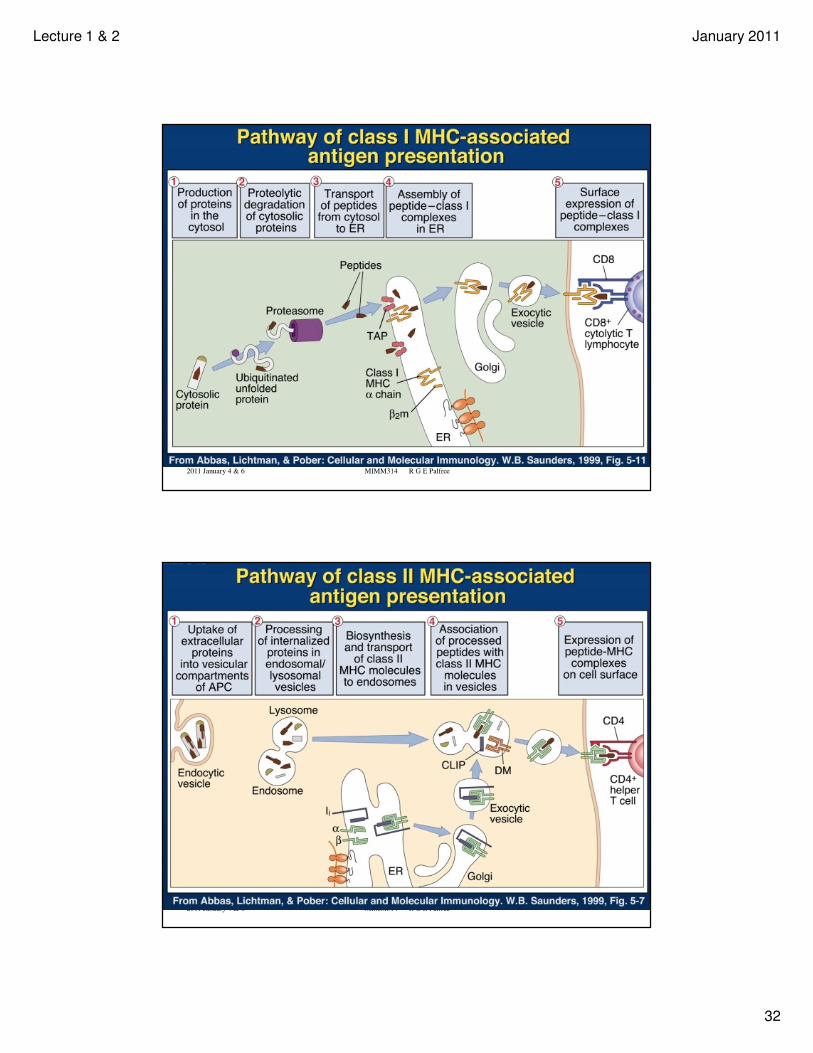
A T cell is naïve until it encounters its antigen presented by a so-called professional antigen-presenting cell (APC). APCs express several different varieties of MHC Class I and II, presenting many different peptides.
In the case of CD4 T cells, the TCR binds to a peptide-MHC class II combination.
The interaction is further stabilized by the T cell CD4 molecule binding to the MHC II molecule on the APC.
Other necessary signals between APC and T cell promote proliferation and differentiation to produce many of the same antigen-specific (and MHC-restricted) effector T cells.
2011 January 4 & 6 MIMM314 R G E Palfree
MHC class I proteins are on the surface of almost all cells. T cells which have the co-receptor CD8 may interact with peptide presented on MHC class I.
MHC class II proteins are limited to so-called professional antigen-presenting cells (APC), which include macrophage, B cells and dendritic cells. T cells which have the co-receptor CD4 may interact with peptide presented on MHC class II.
Dendritic cells are diverse. Like tissue macrophage, monocyte-derived dendritic cells* differentiate in tissues where they become resident. They sample material in their local environment and can be activated by microbial material to become very efficient APCs for activating naïve T cells.
(* Another type of DC, plasmacytoid, has been identified)
Lecture 1 & 2 January 2011
29
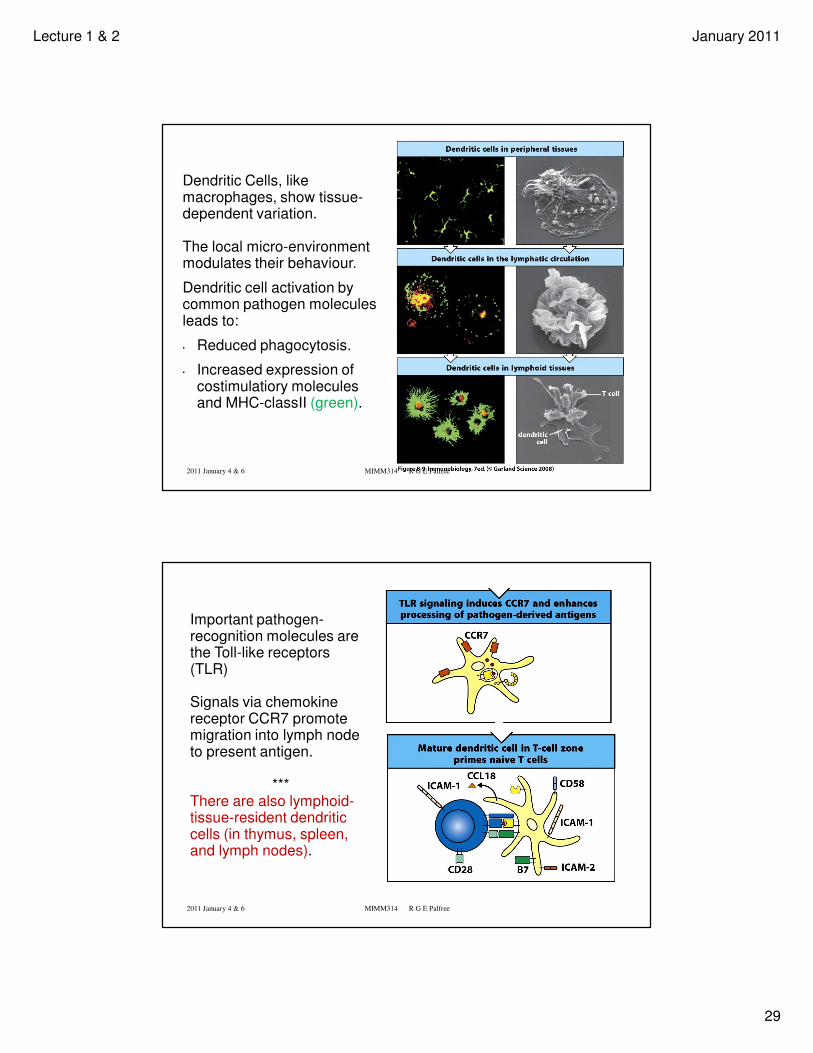
Dendritic Cells, like macrophages, show tissue-dependent variation.
The local micro-environment modulates their behaviour.
Dendritic cell activation by common pathogen molecules leads to:
• Reduced phagocytosis.
• Increased expression of costimulatiory molecules and MHC-classII (green).
2011 January 4 & 6 MIMM314 R G E Palfree
2011 January 4 & 6 MIMM314 R G E Palfree
Important pathogen-recognition molecules are the Toll-like receptors (TLR)
Signals via chemokinereceptor CCR7 promote migration into lymph node to present antigen.
***There are also lymphoid-tissue-resident dendriticcells (in thymus, spleen, and lymph nodes).
Lecture 1 & 2 January 2011
30
2011 January 4 & 6 MIMM314 R G E Palfree
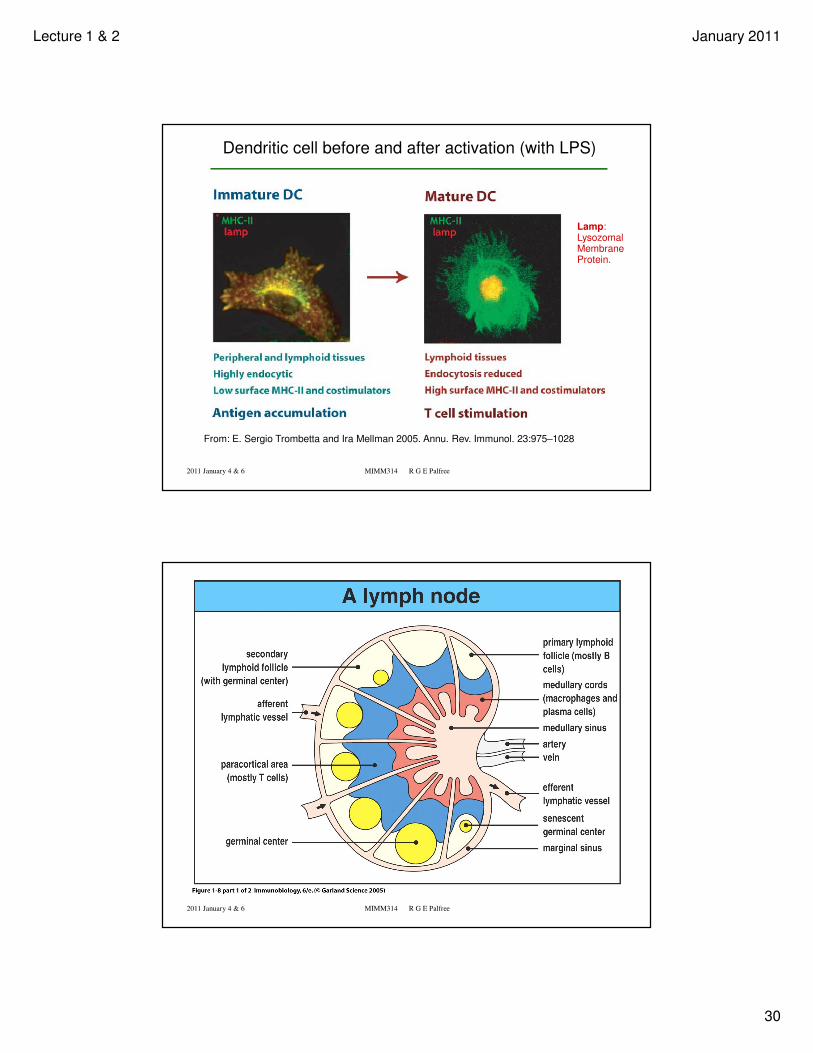
From: E. Sergio Trombetta and Ira Mellman 2005. Annu. Rev. Immunol. 23:975–1028
Lamp:Lysozomal Membrane Protein.
Dendritic cell before and after activation (with LPS)
2011 January 4 & 6 MIMM314 R G E Palfree

Lecture 1 & 2 January 2011
31
2011 January 4 & 6 MIMM314 R G E Palfree 61
Effector T cells then perform several important functions illustrated here.
Each interacts specifically with cells presenting the same peptide-MHCcombination which activated the naïve T cell.
Most B cells require specific help from CD4 T cells to become activated.
They present a set of peptides from any protein within the material which bound to their surface Ig.
2011 January 4 & 6 MIMM314 R G E Palfree
Activated by innate immune responses to pathogen-derived material, or so-called pathogen-associated molecular patterns (PAMPS), Dendritic Cells migrate to secondary lymphoid tissues (e.g. lymph node) and present antigen to T cells. They also express the chemokines to attract naïve T cells, and surface ligands which provide adhesion and costimulation to T cells which bind any of the peptides presented by the APC.
T cells which recognize any of the presented peptides build up a two-way interaction with the APC, proliferate, and differentiate into effector cells which will recognize the same peptide presented by the same MHC molecule on other cells.
The next few slides illustrate the antigen processing pathways and peptide presentation on class I or II MHC.
Lecture 1 & 2 January 2011
32
2011 January 4 & 6 MIMM314 R G E Palfree
2011 January 4 & 6 MIMM314 R G E Palfree

Lecture 1 & 2 January 2011
33
2011 January 4 & 6 MIMM314 R G E Palfree
This is where thymic selection processes are critical.
2011 January 4 & 6 MIMM314 R G E Palfree
Whereas the T cell receptor only binds a polypeptide proteolytic fragment from a protein, and only if it is presented in association with an MHC molecule ...
The B cell receptor (cell surface immunoglobulin), and its secreted form (antibodies), bind to regions (called epitopes) on the surface of native proteins, or many other kinds of molecule including carbohydrate and lipid.
They may bind to epitopes on whole pathogens, aggregates of macromolecules, or even small chemical compounds. (Small chemical antigens are called haptens).
But each antibody is highly specific in its binding.

Lecture 1 & 2 January 2011
34
2011 January 4 & 6 MIMM314 R G E Palfree
When a B cell encounters material which has an epitope for its surface Immunoglobulin (Ig), it binds and internalizes the material.
If there is a protein moiety associated with the epitope, it gets processed into peptides, and some of these are presented by class II MHC molecules on the B cell surface.
If there is an activated T helper type 2 which recognizes any of these peptides, it provides the signals to fully activate the B cell.
2011 January 4 & 6 MIMM314 R G E Palfree
Class I – peptide from intacellular origin.
Class II – peptide from extracellular origin
Class IClass II
Lecture 1 & 2 January 2011
35
2011 January 4 & 6 MIMM314 R G E Palfree
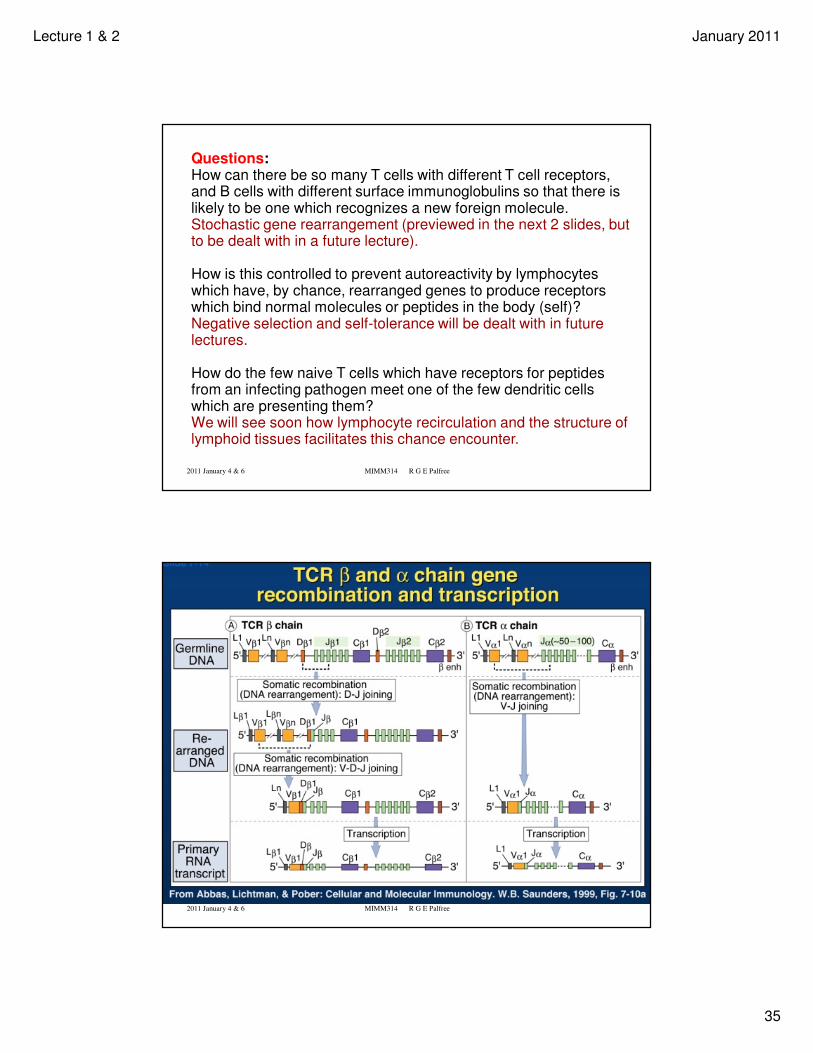
Questions:How can there be so many T cells with different T cell receptors, and B cells with different surface immunoglobulins so that there is likely to be one which recognizes a new foreign molecule.Stochastic gene rearrangement (previewed in the next 2 slides, but to be dealt with in a future lecture).
How is this controlled to prevent autoreactivity by lymphocytes which have, by chance, rearranged genes to produce receptors which bind normal molecules or peptides in the body (self)?Negative selection and self-tolerance will be dealt with in future lectures.
How do the few naive T cells which have receptors for peptides from an infecting pathogen meet one of the few dendritic cells which are presenting them?We will see soon how lymphocyte recirculation and the structure of lymphoid tissues facilitates this chance encounter.
2011 January 4 & 6 MIMM314 R G E Palfree
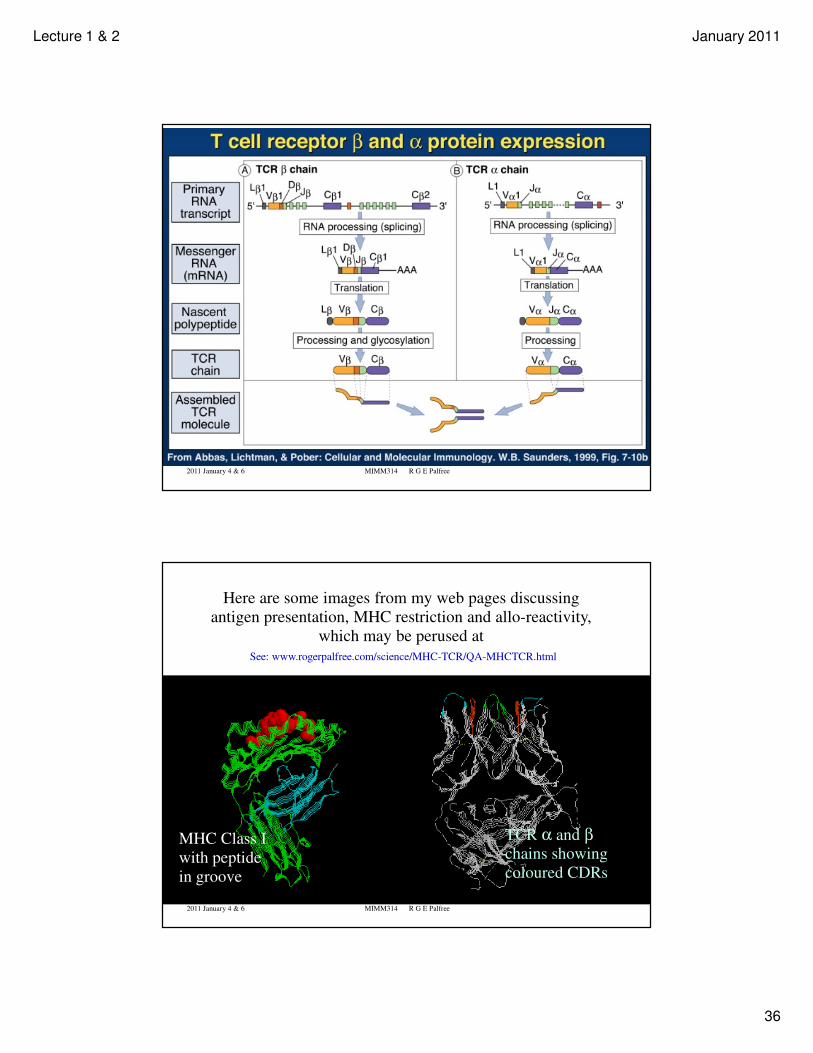
Lecture 1 & 2 January 2011
36
2011 January 4 & 6 MIMM314 R G E Palfree
2011 January 4 & 6 MIMM314 R G E Palfree
T
Here are some images from my web pages discussing
antigen presentation, MHC restriction and allo-reactivity,
which may be perused at
See: www.rogerpalfree.com/science/MHC-TCR/QA-MHCTCR.html
MHC Class I
with peptide
in groove
TCR α and βchains showing
coloured CDRs
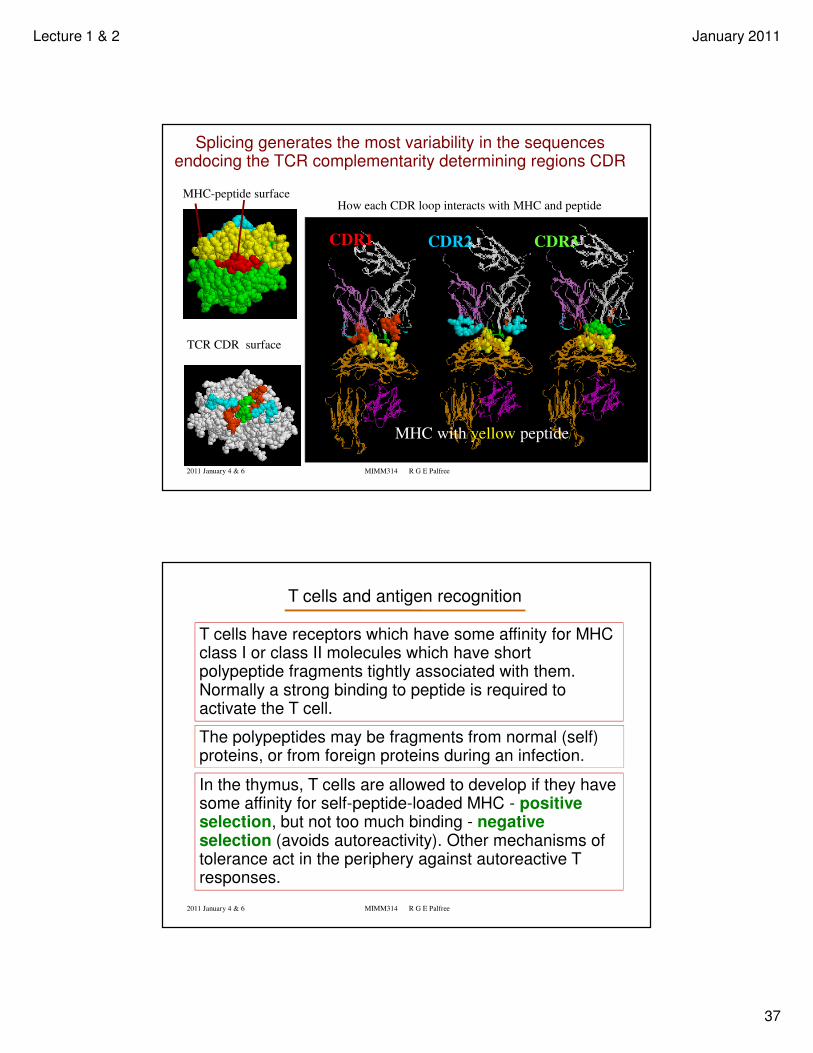
Lecture 1 & 2 January 2011
37
2011 January 4 & 6 MIMM314 R G E Palfree
MHC-peptide surface
TCR CDR surface
How each CDR loop interacts with MHC and peptide
CDR1 CDR2 CDR3
MHC with yellow peptide
Splicing generates the most variability in the sequences endocing the TCR complementarity determining regions CDR
2011 January 4 & 6 MIMM314 R G E Palfree
T cells and antigen recognition
T cells have receptors which have some affinity for MHCclass I or class II molecules which have short polypeptide fragments tightly associated with them. Normally a strong binding to peptide is required to activate the T cell.
The polypeptides may be fragments from normal (self) proteins, or from foreign proteins during an infection.
In the thymus, T cells are allowed to develop if they have some affinity for self-peptide-loaded MHC - positive selection, but not too much binding - negative selection (avoids autoreactivity). Other mechanisms of tolerance act in the periphery against autoreactive T responses.
Related Documents











