DECONTAMINATION OF SPROUTING SEEDS USING HIGH HYDROSTATIC PRESSURE by Swaleha Hudaa Neetoo A dissertation submitted to the Faculty of the University of Delaware in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Animal Science Fall 2011 Copyright 2011 Hudaa Neetoo All Rights Reserved

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DECONTAMINATION OF SPROUTING SEEDS
USING HIGH HYDROSTATIC PRESSURE
by
Swaleha Hudaa Neetoo
A dissertation submitted to the Faculty of the University of Delaware in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Animal Science
Fall 2011
Copyright 2011 Hudaa Neetoo All Rights Reserved

DECONTAMINATION OF SPROUTING SEEDS
USING HIGH HYDROSTATIC PRESSURE
by
Swaleha Hudaa Neetoo
Approved: __________________________________________________________ Jack Gelb Jr., Ph.D. Chair of the Department of Animal and Food Sciences Approved: __________________________________________________________ Robin W. Morgan, Ph.D. Dean of the College of Agriculture and Natural Resources Approved: __________________________________________________________ Debra Hess Norris, M.S. Vice Provost for Graduate and Professional Education

I certify that I have read this dissertation and that in my opinion it meets the academic and professional standard required by the University as a dissertation for the degree of Doctor of Philosophy.
Signed: __________________________________________________________ Haiqiang Chen, Ph.D. Professor in charge of dissertation I certify that I have read this dissertation and that in my opinion it meets
the academic and professional standard required by the University as a dissertation for the degree of Doctor of Philosophy.
Signed: __________________________________________________________ Dallas Hoover, Ph.D. Member of dissertation committee I certify that I have read this dissertation and that in my opinion it meets
the academic and professional standard required by the University as a dissertation for the degree of Doctor of Philosophy.
Signed: __________________________________________________________ Rolf Joerger, Ph.D. Member of dissertation committee I certify that I have read this dissertation and that in my opinion it meets
the academic and professional standard required by the University as a dissertation for the degree of Doctor of Philosophy.
Signed: __________________________________________________________ Kalmia E. Kniel, Ph.D. Member of dissertation committee

I certify that I have read this dissertation and that in my opinion it meets the academic and professional standard required by the University as a dissertation for the degree of Doctor of Philosophy.
Signed: __________________________________________________________ Wallace G. Pill, Ph.D. Member of dissertation committee

ACKNOWLEDGMENTS
I would like to express my sincere appreciation and deep sense of gratitude to my
advisor, Dr. Haiqiang Chen, for his unwavering support and guidance throughout my graduate
study. I especially would like to thank him for granting me much freedom and flexibility in
conducting my research and for his willingness to allow me to explore different avenues. He has
been a great source of encouragement in all of my academic and non-academic endeavors.
Profound thanks to Dr. Dallas Hoover and Dr. Rolf Joerger for all their advice and
guidance throughout my time as a graduate student at University of Delaware. They have always
made themselves very available and accessible to me and have provided constructive comments
at all times. I would also like to thank Dr Kali Kniel for not only serving on my committee but
also for encouraging me to stay on track. I am also deeply indebted to Dr Wallace Pill for his
valuable insight and input through my various projects.
Thanks are also to Dr Thompson Pizzolato, Adrienne Shearer and Dr Larry Cogburn
for helping me in numerous ways in several of my experiments. Special thanks go to all of my
friends, colleagues and acquaintances I have made at the Department of Animal and Food
Sciences at the University of Delaware, for encouraging me to stay positive.

1
TABLE OF CONTENTS
LIST OF FIGURES ........................................................................................................2 LIST OF TABLES..........................................................................................................3 ABSTRACT....................................................................................................................4 INTRODUCTION ..........................................................................................................9 LITERATURE REVIEW .............................................................................................14 POTENTIAL APPLICATION OF HIGH HYDROSTATIC PRESSURE TO ELIMINATE ESCHERICHIA COLI O157:H7 ON ALFALFA SPROUTED SEEDS..................................................................................64 FACTORS AFFECTING THE EFFICACY OF PRESSURE INACTIVATION OF ESCHERICHIA COLI O157:H7 ON ALFALFA SEEDS AND SEED VIABILITY ............................................................91 CONDITIONS FOR ELIMINATION OF ESCHERICHIA COLI O157:H7 ON ALFALFA SEEDS THROUGH A COMBINATION OF HIGH HYDROSTATIC PRESSURE AND MILD HEAT .......................................116 INACTIVATION OF SALMONELLA AND ESCHERICHIA COLI O157:H7 ON ARTIFICIALLY CONTAMINATED ALFALFA SEEDS USING HIGH HYDROSTATIC PRESSURE .......................................................................142 PRE-SOAKING OF SEEDS ENHANCES PRESSURE INACTIVATION OF E. COLI O157:H7 AND SALMONELLA SPP. ON CRIMSON CLOVER, RED CLOVER, RADISH AND BROCCOLI SEEDS .............................................170 INDIVIDUAL AND COMBINED APPLICATION OF DRY HEAT WITH HIGH HYDROSTATIC PRESSURE TO INACTIVATE SALMONELLA AND ESCHERICHIA COLI O157:H7 ON ALFALFA SEEDS ......198

2
LIST OF FIGURES
Figure 2.1 ................................................................................................................12 Figure 2.2 ................................................................................................................39 Figure 2.3 ................................................................................................................41 Figure 3.1 ................................................................................................................81 Figure 3.2 ................................................................................................................81 Figure 4.1 ..............................................................................................................105 Figure 4.2 ..............................................................................................................105 Figure 4.3 ..............................................................................................................106 Figure 5.1 ..............................................................................................................132 Figure 5.2 ..............................................................................................................132 Figure 5.3 ..............................................................................................................133 Figure 5.4 ..............................................................................................................133 Figure 6.1 ..............................................................................................................159 Figure 7.1 ..............................................................................................................182 Figure 7.2 ..............................................................................................................182

3
LIST OF TABLES
Table 2.1 ................................................................................................................21 Table 3.1 ................................................................................................................82 Table 3.2 ................................................................................................................82 Table 3.3 ................................................................................................................83 Table 4.1 ..............................................................................................................106 Table 4.2 ..............................................................................................................107 Table 4.3 ..............................................................................................................107 Table 4.4 ..............................................................................................................108 Table 4.5 ..............................................................................................................108 Table 5.1 ..............................................................................................................134 Table 6.1 ..............................................................................................................160 Table 6.2 ..............................................................................................................160 Table 6.3 ..............................................................................................................161 Table 6.4 ..............................................................................................................161 Table 6.5 ..............................................................................................................161 Table 6.6 ..............................................................................................................162 Table 7.1 ..............................................................................................................193 Table 7.2 ..............................................................................................................193 Table 7.3 ..............................................................................................................194 Table 7.4 ..............................................................................................................195 Table 8.1 ..............................................................................................................220 Table 8.2 ..............................................................................................................221 Table 8.3 ..............................................................................................................221 Table 8.4 ..............................................................................................................222 Table 8.5 ..............................................................................................................223 Table 8.6 ..............................................................................................................224 Table 8.7 ..............................................................................................................225

4
ABSTRACT
Escherichia coli O157:H7 and Salmonella spp. have been associated with seed sprout
outbreaks. The majority of these outbreaks have been traced to sprout seeds contaminated with
low levels of pathogens. Decontaminating sprouting seeds presents a unique challenge in that
even a low residual pathogen population remaining on contaminated seed after treatments
appears capable of growing to very high levels during sprouting. The objective of our study was
to evaluate the potential of using high hydrostatic pressure (HHP) technology alone or in
combination with other hurdles to decontaminate sprouting seeds from Escherichia coli
O157:H7 and/or Salmonella spp.
In our first study, alfalfa seeds were inoculated with a cocktail of five strains of E. coli
O157:H7 and subjected to various combinations of pressure levels and exposure times. When a
pressure level of 650 MPa at 20°C was applied, the threshold time required to achieve complete
elimination was 15 min. Un-inoculated seeds pressure-treated under this condition successfully
sprouted achieving a germination rate identical to untreated seeds after eight days of sprouting.
The second study demonstrated that soaking seeds prior to pressure treatment played a critical
role in enhancing the pressure inactivation of E. coli O157:H7. Seeds soaked in water for 60 min
followed by treatment at 600 MPa for 2 min at 20°C were decontaminated and had a germination
rate of 91% which was 4% lower than that of the untreated seeds (P > 0.05). The third and fourth
study highlighted the effectiveness of high-pressure treatment in combination with mild heat to
eliminate E. coli O157:H7 and Salmonella on artificially contaminated alfalfa seeds. Treatments
carried out at 600 MPa for 2 min at 40°C and 400 MPa for 5 min at 45°C were equally lethal to

5
both pathogens. When both treatments were compared in terms of their impact on the seed
viability, the process of 600 MPa for 2 min at 40°C was the most desirable achieving initial and
final germination percentages of 91 and 96% after 3 and 8 days of sprouting respectively,
compared with 98 and 100% for the untreated control. The applicability of HHP to
decontaminate other types of sprouting seeds was examined in our fifth study. Crimson clover,
red clover, radish and broccoli seeds inoculated with E. coli O157:H7 and Salmonella were
soaked in water prior to pressure treatment. Pre-soaking the different types of seeds in water for
30, 60 or 90 min at ambient or refrigeration temperatures followed by HHP at 600 MPa for 2 or 5
min at 20°C significantly (P < 0.05) enhanced the pressure inactivation of the inoculated
pathogens although it resulted in a variable reduction in their germination rates depending on the
seed type and process conditions. Red clover seeds pre-soaked for 60 min at 4°C prior to HHP at
600 MPa for 5 min attained cumulative germination percentages of 95% after 8 days of
incubation with a slight decrease in their sprouting yield (P > 0.05). Our final study
comparatively assessed the efficacy of dry heat treatments alone or in conjunction with high
hydrostatic pressure (HHP) to eliminate a ~5 log CFU/g load of Salmonella and E. coli O157:H7
on alfalfa seeds. Dry heating alone at 65°C for 10 days or 70°C for 24 h eliminated a ~ 5 log
burden of Salmonella and E. coli O157:H7. Moreover, dry heating at 60 and 65°C for 24 and 12
h respectively followed by HHP at 600 MPa and 35°C for 2 min also eliminated both enteric
pathogens at a variable expense in the sprouting yield.
Taken together, we can ascertain that high pressure processing is an intervention strategy
that can potentially deliver > 5 log reduction and elimination of enterohemorrhagic E. coli
O157:H7 and Salmonella spp. on sprouting seeds with a variable impact on the seed viability

6
indices. The treatment consisting of pre-soaking seeds for ≤ 1 hour at refrigerated temperatures
before pressurization at 600 MPa for 2 min at 20°C offers much commercial scope. Indeed, not
only has this process been demonstrated to successfully decontaminate several types of sprouting
seeds with minimal impact on their vigor, it can also be readily scaled-up from research to high-
volume industrial process scale given the isostatic nature of the pressure process and the ambient
temperature application of the technology.

7
Chapter 1
INTRODUCTION
In many countries worldwide, including the United States, consumption of seed sprouts
has increased in recent decades with the advent of nutraceuticals, phytochemicals (Shetty et al.,
2003) and the shift of consumer preference towards health foods (Gabriel, 2005; Penas et al.,
2008). Alfalfa sprouts are one of the most common sprouts consumed in the United States due to
availability and nutritional value (Fett et al., 2005). However, the sprouting process makes the
commodity susceptible to microbial contamination and growth, thereby compromising the safety
and quality of the sprouts (Gabriel, 2005).
Alfalfa is one of the oldest cultivated plants (Munoz et al., 2007). Originally, it was
called Lucerne, or buffalo herb, and the Arabic named this plant “Alfalfa” which means “Father
of all foods”. The Arabs started eating it after observing its rejuvenating effect on their grazing
animals. Modern research shows that alfalfa also contains a considerable amount of therapeutic
plant compound canavanine, which is an amino acid analogue. Sprouts are also rich in
phytoestrogens, which have diverse functions analogous to those of human estrogen without
their adverse effects. In addition, alfalfa sprouts are also a great source of saponins and active
antioxidants (Gabrovska et al., 2005).
Recently, alfalfa and other types of sprouts that are often consumed raw have been
designated as a special food safety problem by the National Advisory Committee on

8
Microbiological Criteria for Foods (NACMCF) because of the propensity of human pathogens
such as Escherichia coli O157:H7 and Salmonella spp. to rapidly multiply on sprouts during
sprouting and because of lack of a post-harvest lethal step in the processing of sprouts
(NACMCF, 1999). In the past two decade, consumption of raw or lightly cooked seed sprouts
contaminated with Salmonella enterica or Escherichia coli O157 has been responsible for at
least 41 outbreaks of infection in the United States, resulting in over 2300 reported cases of
foodborne illness. Furthermore, contaminated sprouts have been responsible for more multistate
foodborne outbreaks than any other single produce item (FDA, 2004). In most sprout-associated
outbreaks, the source of contamination was the seeds themselves on the basis of direct isolation
and/or epidemiological evidence (Weiss and Hammes, 2005).
Many studies have thus been performed to determine the effectiveness of a wide range of
treatments for reducing the levels of pathogenic Salmonella spp. and E. coli O157:H7 on seeds
and sprouts (Bari et al., 2003). Decontamination by soaking the seeds in different chemical
solutions has been thoroughly investigated. Chemicals used include chlorine, organic acids,
hydrogen peroxide, peroxyacetic acid and ethanol with reported varying degrees of efficacy
(Bari et al, 2008; Jaquette et al., 1996; Munoz et al., 2007; Weiss and Hammes, 2005). It has
been postulated that bacterial cells most likely reside in seed crevices or between the seed coat
and the cotyledon (Charkowski et al., 2001), thus affording protection from chemical
sanitization. Subsequent sprouting conditions may likely provide suitable conditions of
temperature and moisture for the proliferation of surviving cells of E. coli O157:H7 and
Salmonella. These conditions together with nutrients released by sprouting seeds, help low levels

9
of attached pathogens reach dangerous levels on the finished sprouts (Taormina and Beuchat,
1999).
Conventional thermal processing has the potential to ensure food safety and lead to an
extended shelf-life. However, it often leads to detrimental changes in the sensory and nutritional
qualities of the product (NACMCF, 1999). In recent years, there has been a growing interest in
the application of non-thermal processing technologies such as high hydrostatic pressure (HHP)
technology. HHP applied at ambient or slightly elevated temperatures, presents unique
advantages over conventional thermal treatments because of its ability to inactivate a number of
pathogenic and spoilage microorganisms contained in foods without significantly altering their
sensory and nutritional properties (Arroyo et al., 1999). During high-pressure treatment, pressure
transmission is uniform and instantaneous regardless of the sample size and geometry (Knorr,
1995). Hence during isostatic pressure processing, the food product is compressed by uniform
pressure from every direction and returns to its original shape upon pressure release (Olsson,
1995). HHP may be a promising alternative decontamination method for sprouting seeds. Since
pressure acts uniformly and quasi-instantaneously throughout the pressure chamber and on the
food product irrespective of its shape and dimensions, we anticipate that high pressure will act
uniformly on seeds, equally targeting pathogenic bacteria located on the surface of seeds
(crevices or grooves in the seed coat), on the sub-surface such as between the seed coat and the
cotyledons or internalized within the seeds themselves (Mundt and Hinkle, 1976).
Previously, the use of HHP to decontaminate seeds was reported albeit with limited
efficacy (Ariefdjohan et al., 2004; Wuytack et al., 2003). The aim of the current project is to

10
systematically investigate various intrinsic (product-related) as well as extrinsic (process-related)
factors that affect the pressure-inactivation of bacterial pathogens E. coli O157:H7 and/or
Salmonella on alfalfa and other sprouting seeds as well as the viability retention of seeds.
Ultimately, the goal is to delineate and recommend pressure-processing conditions that can be
feasibly applied by large seed distributors to decontaminate seeds prior to shipment to sprout
growers.

11
References
1. Arroyo, G., Sanz, P.D., Prestamo, G., 1999. Response to high-pressure, low-temperature treatment in vegetables: determination of survival rates of microbial populations using flow cytometry and detection of peroxidase activity using confocal microscopy. Journal of Applied Microbiology 86, 544-556. 2. Bari, M. L., Nazuka, E., Sabina, Y., Todoriki, S., Isshiki, K., 2003. Chemical and irradiation treatments for killing Escherichia coli O157:H7 on alfalfa, radish, and mung bean seeds. Journal of Food Protection 66, 767-774. 3. Bari, M. L., Inatsu, Y., Isobe, S., Kawamoto, S., 2008. Hot water treatments to inactivate Escherichia coli O157:H7 and Salmonella in mung bean seeds. Journal of Food Protection 71, 830-834. 4. Charkowski, A. O., Sarreal, C. Z., Mandrell, R. E., 2001. Wrinkled alfalfa seeds harbor more aerobic bacteria and are more difficult to sanitize than smooth seeds. Journal of Food Protection 64, 1292-1298. 5. Fett, W. F., Tortorello, M., Fu, T., 2005. Seed sprouts: the state of microbiological safety, p. 167-219. In: Matthews, K. R. (Ed.), Microbiology of Fresh Produce. ASM Press, Washington, DC. 6. Food and Drug Administration. 19 August 2004, posting date. Note to firms that grow, condition, store, or distribute seed for sprouting and to firms that produce, pack or ship fresh sprouts. [Online.] Food and Drug Administration, Washington, D.C. http://www.cfsan.fda.gov/~dms/prodpla2.html 7. Gabriel, A. A. 2005. Microbial quality of chlorine soaked mung bean seeds and sprouts. Food Science and Technology Research 11, 95-100. 8. Gabrovska D., Paulickova, I., Maskova, E., Fiedlerova, V., Kocurova, K., Pruchova, J. Strohalm J., Houska, M., 2005. Changes in selected vitamins, micro-organisms counts and sensory quality during storage of pressurized sprouted seed of alfalfa (Medicago sativa L.). Czech Journal of Food Science 23, 246-250. 9. Jaquette, C.B., Beuchat, L.R., Mahon, B.E.,1996. Efficacy of chlorine and heat treatment in killing Salmonella stanley inoculated onto alfalfa seeds and growth and survival of the pathogen during sprouting and storage. Applied and Environmental Microbiology 62, 2212-2215. 10. Munoz, M., Ancos, B. D., Sanchez-Moreno, C., Cano, M. P., 2007. Effects of high pressure and mild heat on endogenous microflora and on the inactivation and sublethal injury of

12
Escherichia coli inoculated into fruit juices and vegetable soup. Journal of Food Protection 70, 1587-1593. 11. NACMCF, 1999. Microbiological safety evaluations and recommendations on sprouted seeds. International Journal of Food Microbiology 52, 123-153. 12. Penas, E., Vidal-Valverde, C., Frias, J., Gomez, R., 2008. Application of high-pressure treatment on alfalfa (Medicago sativa) and mung bean (Vigna radiata) seeds to enhance the microbiological safety of their sprouts. Food Control 19, 698-705. 13. Shetty, K. L., Yuan-Tong, L., Mc.Cue, P., G’Labbe, R., Randhir, R., Chia-Yu, H., 2003. Low microbial load sprouts with enhanced antioxidants for astronaut diet. SAE International. 14. Taormina, P. J., Beuchat, L. R., 1999. Comparison of chemical treatments to eliminate enterohemorrhagic Escherichia coli O157:H7 on alfalfa seeds. Journal of Food Protection 62, 318-324. 15. Weiss, A., Hammes, W.P., 2005. Efficacy of heat treatment in the reduction of salmonellae and Escherichia coli O157:H7 on alfalfa, mung bean and radish seeds used for sprout production. European food research and technology 221, 187-191.
.

13
Chapter 2
LITERATURE REVIEW
Characterization of alfalfa and other sprouting seeds varieties
Alfalfa
Alfalfa (Medicago sativa) is a perennial legume (botanical family Leguminosae) mostly
grown for forage production. This legume originated in Central Asia and the Middle East, with
its name derived from the Arabic word for “best fodder”. The seeds are small, kidney-shaped,
yellow, brown or olive green in color, measuring approximately 2 mm x 1mm (Coburn 1907)
and weighing 2-3 mg each. Alfalfa sprouts are the most popular sprouts in the United States.
They have a sweeter, fresher taste when yellow, although they are greened-up to meet consumer
acceptance. The structure of alfalfa seed is illustrated in Figure 2.1.
Figure 2.1 Adapted from Meyer (1999)
Mung Bean
Mung beans (Vigna radiata) often called “Green Grams” or “Chinese bean sprouts”
are thought to originate from Southern Asia. They are only second to alfalfa in popularity for
home growing. These sprouts have been used widely by people in the Far East for centuries. The

14
small, green-colored seeds are available at most natural food stores and Oriental groceries (ISS,
2011)
Clover
Red clover (Trifolium pratense) and crimson clover (Trifolium incarnatum) belong
to the pea family. The clover plant is thought to originate in Europe and has been used for over
2000 years as a cover crop to improve soil quality. Clover sprouts are very similar to alfalfa
sprouts and are sweeter when they are pure yellow (ISS, 2011).
Radish
Radish (Raphanus sativus) belongs to the crucifer or mustard family and has been
described as the “cousin of cabbage, turnip and mustard”. Radish sprouts probably originate in
Eurasia and are fast growing with a spicy, hot flavor. Daikon radish seeds produce tall, lush
green sprouts, while China Rose or Cherry Bell radish sprouts have a tinge of pink (Sellman and
Sellman, 1981).
Broccoli
Broccoli sprouts (Brassica oleracea var. italica) also belong to the crucifer or
mustard or cabbage family and are especially popular because of their delicious “broccoli-like
flavor”. Broccoli sprouts, are very amenable for home sprouting. This cruciferous vegetable
contains compounds called glucosinolates known to prevent certain types of cancers (ISS, 2011).
The Sprouting Process
In addition to the sprouting seed varieties mentioned above, there are even more seed
types currently used for sprouting in commercial operations and at home. Sprouts can be

15
classified as either green or bean sprouts (Fett et al., 2006). Green sprouts such as alfalfa, clover,
broccoli, radish and sunflower typically require exposure to light at some point in the growing
process to allow for chlorophyll development while bean (mung bean and soybean) sprouts are
propagated under continuous dark and thus do not produce chlorophyll. The sprouting yield and
nutritive value of alfalfa and other sprouts are affected by temperature, moisture conditions,
light, cultivar and/or length of sprouting time (Bass et al., 1988).
Commercial sprouting operations in the U.S. are usually indoor facilities and are
usually small in size with less than 10 employees (Fett et al., 2006). Sprouting seeds are first
disinfected before being processed according to recommendations from the International Sprout
Growers Association (ISGA, 2004) and FDA requirements (FDA, 1999a,b). Sprouts are
propagated by growing them hydroponically under dim light at ambient temperature, in flat
trays, close rotating drums or bins, and watering frequently for 4 to 7 days. The growth
temperature can vary depending on the time of the year and geographic location of the sprouting
facility (Beuchat, 1996). Treated potable water is applied through sprinklers at regular intervals.
After harvest, fresh sprouts are thoroughly washed, centrifuged and packed (NACMCF, 1999).
Due to the limited shelf life after harvest, packaged sprouts are distributed to retail outlets either
locally or regionally (Fett et al., 2006).
Sprouts and their nutritional and health benefits
Sprouts, while inexpensive and easy to grow, afford one of the most concentrated but
truly natural sources of vitamins, minerals, enzymes and amino acids (protein) known. They
have long been famous as a natural healthy food by many consumers in the U.S. and elsewhere

16
and are now a staple of health-conscious people in the United States (Meyerowitz, 1999).
Indeed, they are associated with a strong public perception that they are a very healthy product
since sprouts are most often viewed as an “unprocessed” or “natural” food (Wigmore, 1986).
Sprouts are also considered a healthy food because they are low in fat and calories and high in
fiber (Weissinger et al., 2001), with the nutritional content of dry seeds increasing during the
sprouting process (Rajkowski and Thayer, 2001).
Sprouts contain several nutrients of interest, including minerals, proteins and
vitamins such as ascorbic acid, thiamine, riboflavin and niacin (Bass et al., 1988). Vitamin C
(ascorbic acid) is considered an important nutrient in sprouts and on a fresh weight basis, alfalfa
sprouts provide higher amounts of ascorbic acid and iron than other vegetables such as cabbage,
lettuce, carrots (Bass et al., 1988). The health benefits of sprout consumption and prophylactic
abilities of sprouts, in addition to their nutritional benefits, have also been demonstrated by novel
research efforts in several laboratories around the world (Fett et al., 2006). Alfalfa, radish,
broccoli, clover and soybean contain concentrated amounts of phytochemicals (plant
compounds) that confer chemo-protection against development of hypertension and
atherosclerosis (Meyerowitz, 1999). Studies on canavanine, an amino acid analogue in alfalfa,
have shown that it is protective against pancreatic, colon and leukemia cancers. Plant estrogens
which are known to increase bone formation and density and prevent bone breakdown or
osteoporosis are also particularly abundant in sprouts (Meyerowitz, 2007). Moreover, alfalfa
sprouts are one of the richest food sources of another compound called ‘saponins’ (Meyerowitz,
2007). Saponins act to lower the ratio of Low-density lipoproteins (LDL) and Very Low-density

17
lipoproteins (VLDL) to High density lipoproteins (HDL) fats. Animal studies have demonstrated
their protective effect against arteriosclerosis and cardiovascular disease (Meyerowitz, 2007).
Sprouts also contain an abundance of highly active antioxidants that prevent DNA destruction
and protect the human body from the ongoing effects of ageing. As a matter of fact, a study
demonstrated that among numerous tested vegetables, alfalfa sprouts ranked near the top in
antioxidant activity (Meyerowitz, 2007).
Economic significance of sprouts
According to a study conducted by O. B. Hesterman and L. R. Teuber, both at the
University of California at Davis, people in the United States are eating more sprouts than ever
before (Wigmore, 1986). In California, their report states, there was a seventeen-fold increase in
the use of alfalfa sprouts between 1970 and 1979 and more than 10 million pounds of alfalfa
sprouts are produced by California growers annually. In the face of the increased popularity of
sprouts as a healthy food, the North American (United States and Canada) sprouting industry has
expanded rapidly from only a very few commercial growers in 1970 to approximately 475
growers today with a total product value of approximately $250 millions (Price, 1988; Fett,
2005). Beyond North America, the sprouting industry worldwide comprises approximately 850
sprout growing establishments in Japan, 2000 in Europe, 35 in Australia and New Zealand, 3000
in Korea and more than a million in China (Pierre, 2004). On a world-wide basis, sales of sprouts
are generating revenues in the range of $1 billion, with sprouts representing total sales of $250
millions/ year in the United States and Canada from an annual production of 600-million pounds
of sprouts (Pierre, 2004).

18
Microbiological safety of sprouts
Alfalfa sprouts found in retail salad bars are minimally processed and most often
eaten raw or briefly cooked (Weissinger et al., 2001). As is true for many other types of fresh
produce, alfalfa sprouts typically contain high levels of bacteria, some of which may be
pathogenic. The sprouting process itself involves extensive use of water. The sprouting
environment, characterized by high moisture and warm temperatures, creates a favorable
environment for growth and spread of bacteria (Taormina and Beuchat, 1999). Therefore,
contamination of seeds with low levels of pathogens can result in a final product that supports
rapid growth of these organisms (Fett, 2005).
Source of contamination
Sprouting Seeds
Alfalfa seeds may become contaminated from animal waste while growing in the
field. Contaminated irrigation water, run-off water, sewage and improperly composted manure
may also serve as sources of contamination (Mahon et al., 1997). Harvest, transportation, storage
and distribution operations must be considered potential points of contact with pathogens
(Beuchat, 1996).
Sprouting procedures
The sprouts themselves are produced by first soaking the viable seeds in water and then
placing the seeds in a warm, humid environment for an average of 3-7 days to foster germination
and sprout growth. These conditions are supportive of the rapid proliferation of bacteria. If
pathogens are present on or in the seed, sprouting conditions may favor their proliferation.

19
Several studies have indicated that pathogenic bacteria such as salmonellae and E coli O157:H7
present initially on artificially as well as naturally contaminated seeds have the potential to
increase up to 100,000 folds on sprouts grown at 20 to 30°C (Fett, 2005). The majority of growth
of Salmonella and E. coli O157:H7 on sprouting seed occurs during the first 48 hours (Fett,
2005). For sprouts grown from artificially inoculated seed, maximum populations of Salmonella
and E. coli O157:H7 on sprouting seed ranging from 5 to 8 log CFU/g have been reported
(Andrews et al., 1982; Castro-Rosas and Escartin, 2000; Castro-Rosas and Escartin 1999;
Howard and Hutcheson, 2003; Jaquette et al., 1996; Hara-Kudo et al., 1997; Stewart et al., 2001;
Taormina and Beuchat, 1999; Warriner et al., 2003; Barak et al., 2002). However, the maximum
pathogen populations attained during germination and growth of naturally contaminated seed
under commercial practice may be several log units less than that for inoculated seed (Stewart et
al., 2001). These findings therefore support the fact that alfalfa seeds contain sufficient nutrients
to foster microbial growth, and nutrient leaching could promote growth both in the soaking water
and on the nascent sprouts.
In addition to the growth potential of pathogens during the course of sprouting, Barak et
al. (2002) showed that Salmonella serovars can also attach more tightly to surfaces of alfalfa
sprouts than strains of E. coli O157:H7 and the difference in attachment was proposed to partly
explain the greater number of outbreaks of food-borne illnesses due to Salmonella.
Internalization of these bacterial human pathogens in sprouts has also been demonstrated (Itoh et
al., 1998). The mode of entry of bacterial pathogens into plants is not clearly defined, but it is
possibly due to the systemic uptake at the site of injury where lateral roots emerge (Dong et al.,

20
2003; Hallman et al., 1997).
The sprouting environment also increases the likelihood of cross-contamination.
Sprouting involves the use of copious amounts of water. In cases where spot contamination
occurs in one lot of seeds, sprouting water can act as a vector for contaminating the entire batch
(Taormina et al., 1999). The use of contaminated water at the processing facility, poor handling
and poor hygiene can all serve as vectors for contamination (FDA/NACMCF 1999; Rajkowski
and Thayer, 2001).
Incidence of food-borne illness associated with sprouts
Over the past several years, seed sprouts have become a fresh produce item commonly
linked to food-borne illness (FDA/CFSAN, 1999). Seed sprouts are a special concern because as
described above, bacterial pathogens that may be prevalent at very low levels on sprout seeds at
the time of sprouting can multiply to hazardous levels during the 3 to 10 d sprouting process and
can survive through the typical refrigerated shelf life of the products. Also, seed sprouts are
produced as actual agricultural commodities not subject to sanitation requirements because they
are not primarily regarded as foods for human consumption. The potential for sprouts to transmit
pathogenic microbes has been linked especially to the fact that they are usually eaten uncooked
(Piernas and Guiraud, 1998). The first consumer warning about sprouts was issued by the CDC
in 1997 (Powell et al., 2002). CDC and FDA recommended that people at high risk for systemic
infections (i.e. the elderly, young children and immuno-compromised individuals) not eat raw
sprouts.
Outbreaks have been associated primarily with Salmonella serotypes but have also been

21
attributed to Bacillus cereus, E. coli O157:H7, Yersinia enterocolitica and Listeria
monocytogenes (Portnoy et al., 1976; Sivapalasingam et al., 2004). Although not associated with
sprout-related outbreaks, a number of other human pathogens have been isolated from sprouts.
These include Aeromonas caviae and Aeromonas hydrophila (Callister and Agger, 1987;
McMahon and Wilson, 2001), Staphylococcus aureus (Prokopowich and Blank, 1991) and
Klebsiella pneumoniae (Patterson and Woodburn, 1981; Robertson et al., 2002). Since 1990,
consumption of raw or lightly cooked seed sprouts contaminated with Salmonella enterica
serovars or E. coli O157:H7 or O157:NM has been responsible for at least 41 outbreaks of
infection in the United States, resulting in over 2300 reported cases of foodborne illness (Table
2.1). The U.S. Food and Drug Administration estimates that sprout-linked outbreaks account for
40 percent of all food-borne illness associated with produce (Chapman, 2009; 2011). Due to the
widespread nature of the seed distribution systems, some of these outbreaks have had
international impact (Puohiniemi et al., 1997). In addition to outbreaks occurring in the U.S.,
sprout-related outbreaks of food-borne illnesses have been reported in various other countries
including Canada, Japan, Sweden, Denmark, Holland, Finland and the U.K. (NACMCF, 1999;
Taormina et al., 1999). Between May and November 1996, Japan had experienced the largest
recorded outbreak of E. coli O157 infection (De Roever, 1998) through the consumption of raw
radish sprouts served in school lunches.
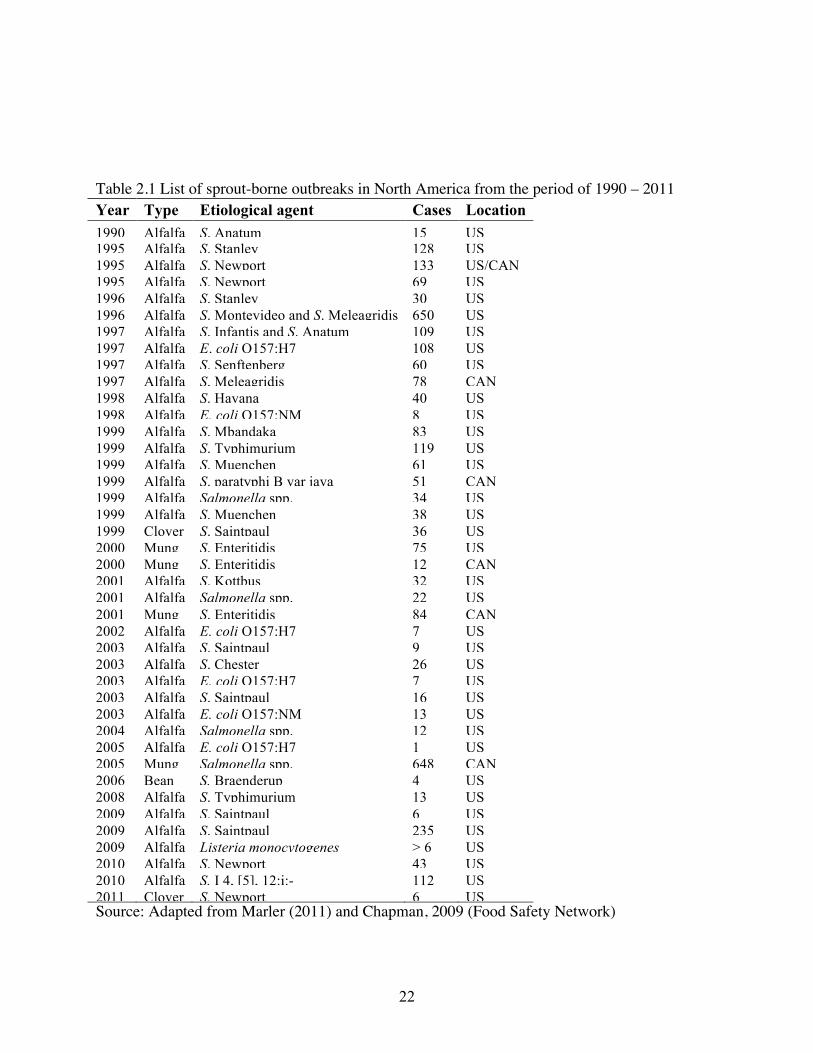
22
Table 2.1 List of sprout-borne outbreaks in North America from the period of 1990 – 2011 Year Type Etiological agent Cases Location 1990 Alfalfa S. Anatum 15 US 1995 Alfalfa S. Stanley 128 US 1995 Alfalfa S. Newport 133 US/CAN 1995 Alfalfa S. Newport 69 US 1996 Alfalfa S. Stanley 30 US 1996 Alfalfa S. Montevideo and S. Meleagridis 650 US 1997 Alfalfa S. Infantis and S. Anatum 109 US 1997 Alfalfa E. coli O157:H7 108 US 1997 Alfalfa S. Senftenberg 60 US 1997 Alfalfa S. Meleagridis 78 CAN 1998 Alfalfa S. Havana 40 US 1998 Alfalfa E. coli O157:NM 8 US 1999 Alfalfa S. Mbandaka 83 US 1999 Alfalfa S. Typhimurium 119 US 1999 Alfalfa S. Muenchen 61 US 1999 Alfalfa S. paratyphi B var java 51 CAN 1999 Alfalfa Salmonella spp. 34 US 1999 Alfalfa S. Muenchen 38 US 1999 Clover S. Saintpaul 36 US 2000 Mung S. Enteritidis 75 US 2000 Mung S. Enteritidis 12 CAN 2001 Alfalfa S. Kottbus 32 US 2001 Alfalfa Salmonella spp. 22 US 2001 Mung S. Enteritidis 84 CAN 2002 Alfalfa E. coli O157:H7 7 US 2003 Alfalfa S. Saintpaul 9 US 2003 Alfalfa S. Chester 26 US 2003 Alfalfa E. coli O157:H7 7 US 2003 Alfalfa S. Saintpaul 16 US 2003 Alfalfa E. coli O157:NM 13 US 2004 Alfalfa Salmonella spp. 12 US 2005 Alfalfa E. coli O157:H7 1 US 2005 Mung Salmonella spp. 648 CAN 2006 Bean S. Braenderup 4 US 2008 Alfalfa S. Typhimurium 13 US 2009 Alfalfa S. Saintpaul 6 US 2009 Alfalfa S. Saintpaul 235 US 2009 Alfalfa Listeria monocytogenes > 6 US 2010 Alfalfa S. Newport 43 US 2010 Alfalfa S. I 4, [5], 12:i:- 112 US 2011 Clover S. Newport 6 US Source: Adapted from Marler (2011) and Chapman, 2009 (Food Safety Network)

23
Organisms of major concern
E. coli O157:H7 and Salmonella are zoonotic pathogens (Tauxe, 1997). The major
reservoir of these pathogens consists of healthy agricultural animals, from which they spread to
an increasingly large variety of food. Traditionally, these pathogens have been mostly associated
with food products of animal origin such as meat and eggs. (Franz et al., 2007) However, recent
outbreaks of food-borne disease associated with the consumption of fresh produce have raised
concern that these products may be effective food vehicles for these enteric pathogens
(Sivapalasingam, 2004; Tauxe, 1997).
Escherichia coli O157:H7
General characteristics E. coli O157 is a Gram-negative, facultative anaerobic (i.e., possesses both respiratory and fermentative metabolic pathways) rod. Most E. coli O157 strains do not ferment
sorbitol and do not contain the enzyme β-glucuronidase (Holt et al. 1994). The optimal growth
temperature for E. coli O157 is 37°C (Holt et al. 1994), but it can survive as low as 7°C and as
high as 46°C. E. coli O157 prefers a neutral pH (6.0-7.0), but it can survive in food at a pH range
of 3.7-4.4 (Weagant and Bound, 2001). Most E. coli O157 strains ferment glucose and some
ferment lactose, producing acid and gas, and are typically oxidase-negative, indole-positive, and
urease negative, and when motile, produce peritrichous flagella (Holt et al. 1994). The unique
characteristics of E. coli O157, such as acid and cold tolerance, make monitoring this foodborne
pathogen critical within the food industry.
There are over 700 antigenic types (serotypes) of E. coli, which are recognized based on

24
the O (somatic), H (flagellar), and K (capsular) antigens (Paton and Paton 1998). There are over
200 O antigens. Flagellar proteins are less heterogeneous than the carbohydrate side chains that
make up the O groups, thus, considerably fewer antigenic types exist (over 30). The O antigens
are composed of lipopolysaccharide complexes, which are part of E. coli cell envelope. It is the
immunogenicity of the polysaccharide repeating units which gives the O antigens their
specificity. The H antigen represents the different flagellin present as part of the flagellar
structure. The K antigen (or the capsular antigen) is mainly an acidic polysaccharide. In general,
the K antigen is left out when serotyping E. coli.
Pathogenesis
E. coli O157:H7, was first identified in 1982 as a causative agent of bloody diarrhea
(hemorrhagic colitis, HC) and hemolytic uremic syndrome (HUS) in humans and associated with
the consumption of undercooked beef. It is considered to be the prototype of EHEC, one of the
six pathotypes of E. coli. Since 1982, the number of E. coli O157:H7 outbreaks and sporadic
cases have been on the rise, and more than 100 outbreaks have been documented. Children under
5 years of age, the elderly and immuno-compromised individuals are in the highest group for
infection by this organism. The infectious dose of E. coli O157:H7 is as low as approximately 10
colony forming units (CFUs) (Kothary and Babu, 2001). Initial symptoms of the infection
include non-bloody diarrhea, “crampy” abdominal pain and short-lived fever. The severity of
infection with E. coli O157:H7 and O157:NM (non-motile) can range from self-limited watery
diarrhea to life-threatening manifestations, such as hemolytic uremic syndrome (HUS) or
thrombocytopenic purpura (TTP). The major illness caused by the antigen is Hemorrhagic

25
colitis, which is characterized by bloody diarrhea, moderate dehydration and acute abdominal
cramps. HUS occurs in all ages but often in children under 10. The associated complications are
pallor, intravascular destruction of red blood cells, depressed platelet counts, lack of urine
formation and renal failure. While the mortality rate of HUS is 3-5%, many survivors suffer
permanent disabilities, such as renal insufficiency and neurological deficits. On the other hand,
TTP commonly occurs in adults. The disease is similar to HUS except that it causes less renal
damage but affects the central nervous system significantly (Doyle, 1991).
The illness-causing Shiga-like toxins (SLT) produced by E. coli O157:H7 is named due
to its similarity to the Shiga toxin produced by Shigella dysenteriae. SLT is a 70 KDa protein
composed of a single A subunit and five B subunits. The B subunit confers tissue specificity,
enabling the toxin to adhere to a specific glycolipid receptor on endothelial cell surfaces. The A
subunit is then delivered to the host cell where it binds with 28S ribosome subunit, inhibits
protein synthesis and kills kidney cells which ultimately leads to HUS. In addition to the SLT,
other virulent factors of E. coli O157:H7 include a chromosomal eaeA gene that encodes for an
outer membrane protein called intimin, which is associated with attachment. Once inside the gut,
EHEC adheres to the intestinal mucosa. A/E (attachment and effacement lesions) are produced in
the brush border microvillus membrane (Knutton et al., 1989). The A/E lesions are initialized
when EHEC injects effector proteins into host cells, which in turn modify the cells function. The
effector proteins bind cytoskeletal proteins and cause them to accumulate at the attachment site,
inducing the attachment and effacement lesions and creating the formation of a pedestal-like
structure (Kodama, 2002). The genes necessary for A/E are encoded on the EHEC chromosome (

26
Sperandio et al., 2006) region, which is highly conserved among intestinal pathogens that
produce A/E lesions (McDaniel et al., 1995).
E. coli O157:H7 also carries a 60 MD plasmid, termed “pO157” or “the large plasmid”.
The functions of this plasmid have not been completely elucidated but it does play a role in
virulence. In addition to virulence genes pO157 also encodes many other genes. These genes
include type II secretion system genes (Tzipori et al., 1987), regulation and expression genes,
replication and maintenance genes, insertion sequences, and some antibiotic resistance genes
(Burland et al., 1998).
Salmonella
General characteristics
Salmonella are Gram negative, facultative anaerobic, rod-shaped, non-sporeforming,
motile bacteria. As a member of the family of Enterobacteriacea, it was named after D.E.
Salmon, a bacteriologist who had identified Salmonella choleraesuis in 1885 (Tauxe, 1991). The
genus encompasses a large taxonomic group with over 2500 serovars under two species,
Salmonella enterica and Salmonella bongori (Wang, 2006), which are classified according to
biochemical characteristics and the immunoreactivity of two surface structures, the O and H
antigens. The O antigen represents a Salmonella specific polysaccharide, and the H antigen
represents the filamentous portion of the bacterial flagella. Variation on these structures results in
different classification within the Salmonella genus. The serotypes commonly isolated from
humans, agricultural products and foods, belong to the subspecies enterica. Salmonella can grow
in temperatures from 2 to 47°C (with the optimum being 25 to 43°C). Most of them can ferment

27
glucose and certain other monosaccharides but generally cannot ferment lactose, sucrose or
salicin. It can grow in the low pH environment of the human stomach, however Salmonella can
be killed under high salt concentrations (Wang, 2006).
Pathogenesis
Salmonella is a well-characterized bacterial food-borne pathogen. Estimates indicate that
Salmonella accounts for approximately 1.0 million illnesses, 19336 hospitalizations and 378
deaths per year in the U.S. resulting in a cost of 4.0 billion dollars annually in medical expenses
and productivity losses (Scallan et al., 2011). The disease caused by Salmonella is called
salmonellosis and the two main manifestations of the disease are typhoid or typhoid-like fever
and gastroenteritis. The clinical syndromes linked to human salmonellosis include enteric fever,
enterocolitis and invasive systemic disease. Enteric fever is caused by Salmonella Typhi and
Salmonella Paratyphi A, B or C (Tauxe, 1991). The main symptoms consist of watery diarrhea,
prolonged and spiking fever, nausea and abdominal cramps. The symptoms of enterocolitis
include severe abdominal pain, diarrhea, vomiting and fever. All serotypes of Salmonella are
potentially pathogenic to humans. The mortality rate of human salmonellosis depends on the age
of the subject, with an average of 4.1% (Wang, 2006). Special populations such as newborns, the
elderly, and patients with immune deficiencies are particularly prone to the infection (D’Aoust,
2001). In general, the infectious dose of Salmonella spp. was thought to be in excess of 10,000
cells but a number of outbreaks have been reported whereby the infectious dose was found to be
very low (10-100 cells), depending on the type of food, strain type, the physiological state of
bacteria and the characteristics of the host (Molbak et al., 2006). The establishment of a human

28
Salmonella infection depends on the ability to survive the environment outside the digestive
system, the ability to survive the gastric acid of the human stomach and the ability of the
pathogen to attach (colonize) and enter (invade) intestinal cells. For the latter, Salmonella must
compete with indigenous gut micro-organisms for suitable attachment sites. Diarrhea associated
with salmonellosis is thought to appear in response to bacterial invasion of intestinal cells rather
than the action of enterotoxins (Molbak et al., 2006). A main difference with other bacterial
intestinal pathogens such as Shigella and E. coli, which replicate within the cytoplasm of host
cells, is that Salmonella is confined to endocytic vacuoles in which bacterial replication takes
place. The infected vacuoles move and release Salmonella cells into the tissue. Prior to invasion
of intestinal cells, Salmonella has to encounter and attach to these cells. This involves several
types of fimbriae or pili. Genes coding for fimbriae are located on the chromosome and open
plasmids. Other virulence factors of Salmonella include siderophores (to retrieve essential iron
from the host) and enterotoxins (Molbak et al., 2006).
It has been estimated that more than 200 virulence factors are responsible for the
pathogenic properties of Salmonella. The virulence determinants are encoded by Salmonella
pathogencity islands or plasmids (Molbak et al., 2006). During the initial stage of salmonellosis,
at least six different adhesion factors are involved in the intestinal colonization of the pathogen.
SPI-1 is responsible for the invasion of Salmonella into the gastro-intestinal epithelium. The
proteins secreted by the bacteria are delivered to the host cells and lead to cytoskeletal
arrangement, membrane ruffling, and bacterial uptake by macro-pinocytosis. Once inside, the
engulfed bacteria reside within and proliferate in the membrane-bound vacuoles. An SPI-2

29
encoded type III secretion system might be responsible for the survival of Salmonella in the
intracellular compartment by avoiding lyzosomal fusion, a host defense mechanism that usually
targets invasive microbes (Hansen-Wester and Hensel, 2001; IFT, 2002). In addition, toxins play
a notable role in the illness. Most serotypes of S. enterica can produce a thermolabile
polypeptide enterotoxin that causes diarrhea. Cytotoxins located in the bacterial outer membrane
might be involved in spreading of Salmonella into deeper host tissues via inhibition of protein
synthesis and lysis of host cells. An endotoxin, also located in the bacterial outer membrane, is
associated with leukocyte-dependent inflammatory response (D’Aoust, 2001).
Sprout industry regulations
In the face of numerous outbreaks and vast number of recalls due to contamination of
seeds and sprouts, several guidelines and regulations have been published. On October 27, 1999,
the FDA and the Center for Food Safety and Applied Nutrition (CFSAN) published a series of
papers as “guidance for the industry” (NACMCF, 1999) entitled “Guidance for Industry:
Reducing Microbial Food Safety Hazards for Sprouted Seeds” and “Guidance for Industry:
Sampling and Microbial Testing of Spent Irrigation Water During Sprout Production” (FDA
1999a,b).These guidances are intended to provide recommendations to suppliers of seed for
sprouting and sprout producers about how to reduce microbial food safety hazards common to
the production of raw sprouts to ensure that sprouts are not a cause of foodborne illness and to
ensure that they comply with the food safety provisions of the Federal Food, Drug, and Cosmetic
Act. The first guidance is based largely on recommendations from the National Advisory
Committee for Microbiological Criteria for Food's report entitled “Microbial Safety Evaluations

30
and Recommendations on Sprouted Seeds'' (NACMCF, 1999). This document identifies the
preventive controls that the FDA believes should be taken immediately to reduce the public
health risks associated with raw sprouts and to ensure that sprouts are not adulterated under the
food safety provisions of the Food, Drug and Cosmetic Act (FDCA). The failure to adopt
effective preventive methods of control assumes that the product was prepared under unsanitary
conditions, which may render it injurious to health. Food produced under such conditions is
adulterated under the act [21 U.S.C. 342 (a) (4)]. The FDA would enforce actions against any
party which does not have effective preventive controls in place, particularly, microbial testing.
This paper also provides guidelines and recommends good agricultural practices (GAPs) for seed
production, conditioning, storage and transportation and good manufacturing practices for sprout
production. It also advocates seed treatment with an approved antimicrobial (i.e. use of the
recommended 20,000 ppm calcium hypochlorite soak), testing for pathogens in the spent
irrigation water, and implementation of traceback systems (FDA, 1999a). The second guidance is
intended to assist sprouters in implementing one of the principal recommendations i.e., microbial
testing. This guidance is designed to assist sprouters in designing a microbial testing program to
ensure adulterated product does not enter commerce. Specifically, this guidance recommends
testing spent irrigation water from each individual batch or production lot of sprouts for two
pathogens, E. coli O157:H7 and Salmonella. The microbial testing guidance also provides
instructions for the sampling and testing of sprouts for those instances when it is not possible to
test spent irrigation water. However, sprouts should not be tested in lieu of irrigation water when
spent irrigation water is available (FDA, 1999b).

31
Global approach for reduction of risks of seed contamination
Various measures should be taken to prevent the initial contamination of seeds or to
eliminate possible contaminants. This can be achieved by the application of good food hygiene
practices, Good Agricultural Practices (GAPs), Good Manufacturing Practices (GMPs) and
Hazard Analysis Critical Control Points (HACCP) at all stages of seed and sprout production
including growth, harvesting and distribution of the seeds (raw material), and germination and
handling of the sprouts (finished products). Microbial contamination of seeds and sprouts can
occur at various stages, hence control methods must be devised to address the potential vehicles,
vectors or routes of contamination, which pose the greatest risks to the final sprout product.
Moreover, research on mitigating the risks from sprouted seeds using traditional and novel seed
decontamination strategies is also patent.
GAPs
Farmers and seed producers must undertake a risk assessment, and ensure control by
application of GAP principles and sourcing seeds from areas where they are not likely to come
into contact with human/animal waste (Beales, 2004). It is important that seed producers from all
countries differentiate between seeds intended for sprouting for human consumption and those
used for animal feed and are aware of the microbiological risks associated with this type of
product. Particular consideration should be paid to the presence of animal production facilities
and related factors that could lead to the crop being exposed to untreated manure, the presence of
domestic grazing animals on fields intended for seed production and the sources of agricultural
water used.

32
GMP and HACCP
In addition to contamination of seeds during production, seeds destined for sprouting can
also be contaminated during post-harvest operations such as transport, conditioning, grading,
storage and packaging. It is thus essential to follow GMP during seed handling steps such as seed
cleaning and storage as well as during downstream sprouting procedures such as sprout
production, packaging and distribution.
HACCP describes a system of control for assuring food safety and provides a more
proactive and structured approach to the control of identified hazards than that achieved by
traditional inspection and quality control procedures. It has the potential to identify areas of
concern where failure has not yet been experienced, making it particularly useful for new
operations. The International Sprout Growers Association (ISGA, 2004) has produced a HACCP
checklist, which details critical control points during the production of sprouted seeds, the
potential hazards and the control measures that can be taken (Beales, 2004).
Methods for seed decontamination
There have been more reported studies on interventions for the decontamination of
sprouting seeds and sprouts than from any other type of produce (Fett, 2005) Since seeds are
thought to be the primary source of pathogens for sprout-related food-borne outbreaks, most
potential interventions have been tested on seeds rather than on sprouts themselves (Fett, 2005).
Adequate decontamination of seeds presents a unique challenge due to the ability of surviving
bacteria to grow rapidly during the sprouting process. Hence the goal should be to completely
eradicate any pathogens present on seeds. This must be accomplished while maintaining post-

33
treatment seed viability as well as a commercially acceptable sprout yield, appearance and shelf-
life (Fett et al., 2006).
Chemical Interventions
In 1999, the FDA published guidance documents recommending that commercial sprout
growers treat sprout seeds with one or more antibacterial treatments such as 20,000 ppm of
calcium hypochlorite that have been approved for reduction of pathogens on seeds or sprouts,
with at least one approved antimicrobial treatment applied immediately before sprouting (FDA,
1999a). Population reductions reported after treatment of alfalfa seed artificially inoculated with
Salmonella or E. coli O157:H7 using 16,000 to 20,000 ppm of available chlorine has varied
considerably among different laboratories ranging from 2 to 4 log10 CFU/g (Fett, 2005). One
consistent finding among the various laboratories is that the two pathogens on artificially
inoculated sprout seeds are not completely eliminated even by treatment with up to 20,000 ppm
of available chlorine for 10 to 15 minutes (Fett, 2005). The inconsistent population reduction
results across different research laboratories may be due to several factors including differences
in the initial level of contamination or differences in the location of the pathogens on the
contaminated seed such as the surface, sub-surface or internal tissues of seeds (Charkowski et al.,
2001).
Numerous chemical treatments in addition to chlorine treatments have been tested
individually or in combination for pathogen reduction on seeds. Chemicals tested have included
aqueous acetic acid, calcinated calcium, carvacrol, cinnamic aldehyde, citric acid, Citricidal,
CitroBio, ethanol, eugenol, linalool, methyl jasmonate, sodium carbonate, thymol, trisodium

34
phosphate, Tsunami 2000, Tween 80, Vegi-clean and Vortex (Fett, 2005). Treating with aqueous
chemicals at elevated temperatures can lead to greater reductions of pathogen populations on
seed, but is often detrimental to seed germination (Scouten and Beuchat, 2002). Addition of high
levels of the surfactant Tween 80 (1% w/v) to 1% Ca(OH)2 led to only an additional 1 log10
CFU/g or less in the population of Salmonella on alfalfa seed. Sonication of seed during
treatment with aqueous chemicals antimicrobial compounds also did not have a significant
effect, only slightly increasing the log kill obtained. Treatment with gaseous acetic acid was
reported to eliminate both Salmonella and E. coli O157:H7, but not Listeria monocytogenes,
from artificially inoculated mung bean seed without reducing seed germination (Delaquis et al.,
1999). However, similar treatments of inoculated alfalfa seed led to either unacceptable
reductions of seed germination or were not effective (Delaquis et al., 1999).
Physical Interventions
Enomoto et al. (2002) previously studied the ability of hot-water treatments to eliminate
E coli ATCC 25922 from inoculated alfalfa seeds and reported a > 4 log reduction with no
significant reduction in the germination rate. Hot-water treatments (57-60°C for 5 min) of
artificially contaminated alfalfa seeds revealed > 2.5 log CFU/g reduction in the population of S.
enterica (Jaquette et al., 1996). However, slightly higher temperatures or longer treatment times
brought about a dramatic decrease in seed viability (Fett et al., 2006). Other authors have
reported that thermal treatments of sprouting seeds have not delivered consistent results due to
lowered effectiveness and/or detrimental effects on seed germination (Fett et al., 2006).
Moreover, with thermal treatments, there is often a very narrow time-temperature regime where

35
pathogen levels can be effectively reduced or eliminated whilst maintaining the seed germination
rate and sprouting yield at an acceptable level. As a result, scaling up of hot-water treatments for
large-volume processing may be problematic.
Several other physical treatments have also been tested for sanitizing sprout seed. In
2000, the FDA approved the application of ionizing radiation at doses up to 8 kGy on sprout
seeds (FDA, 2000). Treatment with ionizing radiation has been shown to significantly reduce
bacterial pathogens on sprout seed: exposure of inoculated alfalfa seed to a 2 kGy dose of
gamma radiation led to a 3.3 and 2.0 log reduction in E. coli O157:H7 and Salmonella
populations, respectively, while still maintaining acceptable sprout yields (Thayer et al., 2003;
Rajkowski and Thayer, 2001; Fan et al., 2004). Higher doses led to commercially unacceptable
reductions in yields. For alfalfa seed naturally contaminated with Salmonella and treated with
gamma irradiation, Thayer et al. (2003) reported a D-value of 0.81 kGy. An absorbed dose of 4
kGy was required to eliminate the pathogen, a dosage that resulted in significant reductions in
yield. Electron bean radiation (low energy electron beam, energies < 300 kV) may also be useful
for reducing pathogen populations on the surface of seed, but may have lowered penetration
ability compared with gamma radiation.
In addition to stand-alone treatments, various treatment combinations (hurdle concept)
for reducing the contamination on sprout seed have been tested. Bari et al. (2003) reported that
the sequential application of dry heat followed by treatment with hot acidic electrolyzed
oxidizing (EO) water and sonication was able to reduce populations of E. coli O157:H7 on
artificially inoculated mung bean seed by 4.6 log but the combination treatment was less

36
effective when tested against inoculated radish and alfalfa seed. Seed germination and
subsequent sprout growth were not adversely affected. In the same study, a dry heat (50°C, 1
hour) seed treatment in combination with exposure to 2 to 2.5 kGy of gamma radiation led to the
elimination of the pathogen on mung bean, radish and alfalfa seed, but resulted in a reduction in
the yield, most significantly for mung bean and radish. Sharma (2002) found that treating alfalfa
seed inoculated with E. coli O157:H7 first with ozone (continuous sparging in water) followed
by a dry heat treatment (60°C, 3 hours) led to a greater than 4 log reduction of the pathogen
population although survivors were still detected by enrichment.
Biological interventions
Unfortunately, there is little published information on the application of competitive
exclusion to control human pathogens on produce. Biopreservation can be challenging since the
ideal biocontrol culture for use on sprouting seeds and sprouts would consist of a nonpathogenic
microorganism that is not likely to undergo mutations, can easily be propagated using low cost
substrates, has a long shelf-life, can be easily administered to seeds and/or sprouts, is highly
effective on a variety of sprout types and against several human pathogens and represents a low-
cost intervention for the grower (Fett et al., 2006).
Most of the interventions on biological control of bacterial human pathogens on produce
have examined the use of lactic acid bacteria (LAB) as antagonists. LAB are attractive options
for commercial biological control agents due to their ubiquity on sprout surfaces (Patterson and
Woodburn, 1981; Cai et al., 1997), their ability to produce multiple antimicrobial agents
including bacteriocins, hydrogen peroxide and organic acids in vitro, their wide acceptance in the

37
food industry for fermentation and their lack of known pathogenicity. A strain of Lactococcus
lactis shown to be inhibitory in vitro against Listeria monocytogenes due to acid production was
tested for control of the pathogen on alfalfa seed before sprouting (Palmai and Buchanan, 2002).
Results indicated that the strain was much less inhibitory towards the pathogen in situ than in
vitro, reducing pathogen populations by only less than 1 log.
There is also evidence that competitive exclusion using pseudomonads may constitute a
viable intervention against Salmonella outgrowth on sprouts as shown by Matos and Garland
(2005). Addition of Pseudomonas fluorescens 2-79 to the seed soaking solution at 8 log CFU/ml
led to a 4 log reduction in the population of Salmonella after 1 and 3 days of sprouting of
artificially inoculated alfalfa seeds. There was no adverse effect of inoculation with 2-79 on
sprout appearance.
Another biological approach to controlling pathogen is the application of phage. The use
of lytic phages for curbing bacterial plant pathogens is an active area of research. Pao et al.
(2008) tested the abilities of two lytic phages to inhibit outgrowth of S. enterica on sprouting
broccoli and radish seeds. Application of phage at approximately 6 to 7 log PFU/ml at the time
of germination was not highly effective (less than 1.5 log reduction after 24 h) against
susceptible S. enterica serovars. The host specificity of lytic phage would necessitate the use of
phage cocktails under commercial practice (Pao et al., 2008).
Despite considerable research efforts towards the development of sprout seed sanitizing
methods, there is still a need for highly effective and environmentally friendly seed-sanitizing
strategies that can deliver consistent kill of sprouting seed-borne human pathogens. The use of

38
20,000 ppm calcium hypochlorite presents worker and environmental safety concerns, and may
not always be effective in eliminating human pathogens from contaminated seed lots under
commercial practice, and can be highly detrimental to the germination capability of some seed
types.
There has been limited research on the use of nonthermal processing methods such as
high hydrostatic pressure (HHP) for reducing the microbial load on seeds (Ariefdjohan, 2004;
Penas et al., 2008; Wuytack, 2003) and sprouts (Munoz et al., 2007). The latter researchers have
either reported a limited decontamination efficacy of HHP on seeds or adverse collateral effects
on the seeds’ germinability. In the current study, the application of HHP to decontaminate alfalfa
and other sprouting seeds is systematically investigated, with due attention paid to treatments of
the seeds prior to HHP or optimization of the HHP conditions to enhance pressure-inactivation of
E. coli O157:H7 and Salmonella with minimal decrease in seed viability.
High Hydrostatic Pressure (HHP)
High Hydrostatic Pressure (HHP), High Pressure Processing (HHP) and Ultra High
Pressure (UHP) are all names for the same process. HHP is a cold pasteurization method
employed for shelf life extension and pathogen reduction while retaining the food’s inherent
color, flavor, nutrients and texture. HHP can inactivate vegetative microbial cells by breaking
non-covalent bonds and causing damage to the cell membrane. High pressure disrupts secondary
and tertiary structures of macromolecules such as proteins and polysaccharides, and alters their
structural and functional integrity in a pressure-dependent way. The process causes non-covalent
bonds to break, interrupting cellular function and inactivating the pathogenic bacterial cells. The

39
product can be solid or liquid food, packaged or not, to which 100-1000 MPa of pressure is
applied at temperatures ranging from 0-110°C, from a millisecond pulse to over 20 min, in batch
or semi-continuous systems. The heat generated during compression adiabatically raises the
temperature of the product about 3°C for every 100 MPa, a change reversed during
decompression (Palou et al., 1999).
Past and present progress in the application of HHP on food products
Studies of the effects of high pressures on foods date back over a century. In 1899, Bert
Hite of the Agricultural Research Station in Morganstown, West Virginia, USA, designed and
constructed a high-pressure unit to pasteurize milk and other food products (Hite, 1899). Hite
constructed a machine that could reach pressures in excess of about 6800 atmospheres
(approximately 700 MPa) and he and his co-workers examined the potential use of HHP
processing for a wide range of foods and beverages, including the pressure inactivation of
viruses. The level of sophistication that was accomplished is remarkable, given the
technological limitations faced during that time period, especially with regard to processing
systems and packaging materials (Hoover, 1993). In 1899, Hite reported that treatment at
pressures of 450 MPa or greater could improve the keeping quality of milk (Hite, 1899). In 1914,
he showed that yeasts and lactic acid bacteria associated with sweet, ripe fruit were more
susceptible to pressure than other organisms especially spore-forming bacteria associated with
vegetables. Compared to today’s HHP processing equipment, the prototype system utilized in the
1890’s by Hite was very primitive. Today, with advances in computational stress analysis and
new materials, high capacity pressure systems can be manufactured to allow reliable HHP

40
treatment of food products at even higher pressures (Hoover, 1993).
Although the potential for HHP processing of foods has been discovered since the late
nineteenth century, its potential and value in food safety and preservation have only been truly
appreciated in the last 30 years. High hydrostatic pressure presents unique advantages over
conventional thermal methods for food processing. Its application at low temperatures permits
the retention of food quality (Knorr 1993). It does not cause environmental pollution and
eliminates the use of chemical additives in food products (Kadharmeston, 1998). Considerable
research and development in HHP processing of foodstuff has taken place worldwide, with a
number of products being successfully commercialized. In 1990, the first high-pressure
processed product, a high acid jam, was introduced to the Japanese retail market. The jams were
vivid and natural in color and taste. In 1991, yogurt, fruit jellies, salad dressings and fruit
products were also made commercially available. In the U.S. and Europe, research and
commercial developments are also continually being made in fruit products, ready meals, dairy
products, condiments, cooked meats and seafood (Rastogi et al., 2007).
Theoretical principles of the HHP process
Two principles describe the effect of HHP. Firstly, the principle of Le Chatelier,
according to which any phenomenon (phase transition, chemical reaction, change in molecular
configuration) accompanied by a decrease in volume can be enhanced by pressure (Palou et al.,
1999). Secondly, the Isostatic rule states that pressure is instantaneously and uniformly
transmitted throughout a sample under pressure, whether the sample is in direct contact with the
pressure medium or hermetically sealed in a flexible package that transmits pressure. In other

41
words, pressure is transmitted in a uniform (isostatic) and quasi-instantaneous manner
throughout the sample and the time necessary for pressure processing is independent of sample
size, in contrast to thermal processing (Hogan et al., 2005). The principle of isostatic processing
is presented in Figure 2.2.
Figure 2.2 (Source: Palou et al., 1999)
Working principles of the HHP process
In a HHP process, the food product to be treated is placed in a pressure vessel capable of
sustaining the required pressure; the product is submerged in a liquid, which acts as the pressure-
transmitting medium. Water may be used as the pressure-transmitting medium, but media
containing castor oil, silicone oil, sodium benzoate, ethanol or glycol can also be used
(Yaldagard et al., 2008). The ability of the pressure-transmitting fluid to protect the inner vessel
surface from corrosion, the specific HHP system to be used, the process temperature range and
the viscosity of the fluid under pressure are some of the factors involved in selecting the medium
(Hogan et al., 2005).
Industrial HHP treatment is currently a batch or semi-continuous process. The selection
of equipment depends on the kind of food product to be processed. Solid food products or foods

42
with large solid particles can only be treated in a batch mode. Liquids, slurries and other
pumpable products have the additional option of semi-continuous production (Ting and
Marshall, 2002). Currently, most HHP machines in industrial use for food processing are batch
systems, whereby the product is placed in a high pressure chamber and the vessel is closed, filled
with pressure-transmitting medium and either pressurized either by pumping medium into the
vessel or by reducing the volume of the pressure chamber, for example using a piston. If water is
used as the pressurizing medium, its compressibility must be accounted for; water is compressed
by up to 15 per cent of volume at pressures above 600 MPa (Palou et al., 1999). Once the desired
pressure is reached, the pump or piston is stopped, the valves are closed and the pressure is
maintained without further energy input. After the required holding time has elapsed, the system
is depressurized, the vessel opened and the product unloaded. The system is reloaded with
product, either by operators or machines depending on the degree of automation (Ting and
Marshall, 2002). High pressure can be generated by direct or indirect compression or by heating
the pressure medium. Most industrial isostatic presses use the indirect compression system.
Indirect compression is achieved using a high-pressure intensifier to pump the pressure medium
from a reservoir into a closed high-pressure vessel until the desired pressure is reached (Figure
2.3.).

43
Figure 2.3 (Source: Mertens 1995)
The total time for pressurization, holding and depressurization is referred as the “cycle
time”. The “cycle time” and the loading factor (i.e. the percentage of the vessel volume actually
used for holding packaged product, primarily a factor of package shape) determined the
throughput of the system. In a commercial situation, with this sort of batch process, a short
holding time under pressure is desirable in order to maximize the throughput of the product.
For any HHP system, the working pressure is a very important parameter, not only
because the initial price of the equipment increases significantly with its maximum working
pressure, but also because a decrease in working pressure can significantly reduce the number of
failures, thereby increasing the working life of the equipment (Otero, 2000). Keeping the sample
under pressure for extended periods of time does not require any additional energy. The work of
compression during HHP treatment will increase the temperature of foods through adiabatic
heating by approximately 3°C per 100 MPa, depending on the composition of the food.

44
Use of HHP to improve food safety and quality
Application of HHP as a method for microbial inactivation has stimulated considerable
interest in the food industry. Currently, research in this area has concentrated mainly on the
effect of HHP on vegetative cells and spores of different pathogenic and spoilage bacterial
species. Over the last decade, consumer demand has increasingly required processed foods to
have more natural flavor and color with a shelf-life that is sufficient for distribution and a
reasonable period of home storage before consumption. This can be achieved by minimal
processing methods that preserve foods but also retain to a greater extent their nutritional quality
and sensory characteristics by using alternative hurdles other than heat as the main preservative
action. HHP is such a minimal processing method that does not result in any substantial increase
in product temperature. There is therefore little damage to pigments, flavor compounds or
vitamins and in contrast to heat processing, the sensory characteristics and nutritional value of
foods are not degraded to a significant extent. The resulting products have higher quality and
consumer appeal in markets where the retention of nutritional sensory characteristics can
command premium price (Fellows, 2000).
Microbial inactivation by HHP
Pressure treatment of microorganisms induces various changes to microbial cellular
targets including changes to the cell permeability and morphology, inhibition of biochemical
reactions essential for cell maintenance and survival, reproduction of the cell and genetic
mechanisms (Patterson, 2005). Although the intricate mechanistic details of microbial
inactivation still remain uncertain, several theories have been proposed over the years in an

45
attempt to shed light on the mechanism of microbial inactivation. To that effect, the different
cellular targets of high pressure involved in bacterial inactivation are described in the following
sections.
Effect of HHP on bacterial cell membrane
The membrane is generally recognized as the main site of pressure damage in
microorganisms as high pressure treatment almost always leads to a perturbation of bacterial
membranes (Patterson, 2005). It is thought that the mechanism of pressure-induced membrane
damage is due to the compression of the fatty acyl chains of membrane phospholipids, causing
them to transition from a liquid crystalline to a gel state resulting in a subsequent loss of
membrane integrity and membrane permeability (Patterson et al., 2007). HHP can also alter other
membrane functions such as active transport or passive diffusion, offsetting the physicochemical
and biochemical homeostatic mechanisms of the cell. Microbial inactivation is also partly due to
loss of functionality of membrane-bound enzymes: membranes of pressure-treated cells were
shown to detach from the surrounding lipid-bilayers, which are responsible for creating an
optimum environment for the activity of membrane-bound enzymes (Yaldagard et al., 2008).
Effect of HHP on bacterial cell morphology
Internal cellular damage has been observed in high-pressure exposed cells viewed under
scanning electron microscopy (Patterson, 2005). Some authors observed the formation of bud
scars on the surface of the cell of Listeria monocytogenes after a 10-min pressure treatment at
400 MPa (Manas and Mackey, 2004). Park et al. (2001) exposed cells of Lactobacillus
viridescens to a pressure level of 400 MPa for 5 min at 25°C and showed that the ultra-structure

46
of pressure-treated cells revealed the presence of nodes on the cell wall. Smelt (1998) stated that
high pressure resulted in the destabilization of ribosomes, causing the number of functional
ribosomes to irreversibly fall below a minimum threshold level required to recover from
sublethal stress, ultimately leading to cell death. Certain authors also observed the extensive
condensation of the nucleoid which became confined to the periphery of the cell membrane
(Manas and Mackey, 2004).
Effect of HHP on biochemical reactions
Several authors reported that pressure affects cellular reaction systems in two apparent
ways: (1) by altering intra-molecular structures and reducing the available molecular space and
(2) by increasing inter-chain reactions at enzyme/substrate interfaces (Hoover, 1993). High-
pressure treatment favors biochemical phenomena that lead to an overall volume decrease while
reactions that result in a volume increase are inhibited. Exposure to high pressure may activate or
inactivate bacterial enzymes as a result of their inherent structural differences as well as their
differential ability to withstand the pressure stress. Pressure effects on enzymes are dependent on
the pressure magnitude: pressure exceeding 300 MPa causes irreversible denaturation while
lower pressure ranging from 100-300 MPa results in reversible structural changes. Enzyme
activation can also occur following disruption of the membranes of sub-cellular compartments in
which the enzymes are usually sequestered, thus facilitating enzyme-substrate interaction.
Inhibitory effects of high pressure on microorganisms can also be caused by enzyme inactivation
(Patterson et al., 2007). Pressurization at room temperature may result in reversible or
irreversible, partial or complete enzymatic activity loss as a result of extensive conformational

47
changes. The severity of those changes will depend on the molecular structure of the enzyme
(tertiary and quaternary structures), conditions affecting the microenvironment of the enzyme
(e.g. pH), pressure level, temperature and pressure exposure time (Palou et al., 1999).
Effect of HHP on cellular proteins
Bartlett (2002) demonstrated that pressure applied at a low level of 53 MPa was able to
induce the expression of a specific class of proteins normally switched on at elevated
temperatures. HHP can also bring about the denaturation of proteins (Heremans, 1995), as
exemplified by the denaturation of lactate dehydrogenase and glyceraldehyde-3-phosphate
dehydrogenase from rabbit muscle and Saccharomyces cerevisiae cells following pressure
application at a level of 200 and 100 MPa, respectively. Protein denaturation induced by HHP is
thought to be one of the causes of bacterial cell injury and/or death. HHP has a greater
detrimental effect on the tertiary and quaternary structures of proteins, making multimeric
enzymes i.e. enzymes with several subunits, more susceptible to HHP than monomeric proteins
(Palou et al., 1999). During HHP, trans-membrane and peripheral membrane proteins can also
become detached following significant perturbation of the membrane during HHP (Patterson,
2005). Moreover, the disruption of membrane-bound enzymes critical for homeostatic regulation
of bacterial cells has also been demonstrated (Manas and Mackey, 2004).
Effect of HHP on genetic mechanisms
Genetic molecules such as nucleic acids (DNA and RNA) are very piezotolerant. The DNA
double helix itself is stabilized by H-bonding and since HHP does not have any negative effect
on H-bond, its structure is stable under pressure. Although DNA is structurally unaffected, its

48
functionality is indirectly affected due to the effect of HHP on enzyme-mediated genetic
processes such as DNA replication and gene transcription (Bartlett, 2002). In E. coli, arrest of
DNA synthesis was observed following application of pressure at a level of 50 MPa. Bartlett
(2002) speculated that the role of membrane fluidity is central to DNA replication, and certain
genetic processes such as replication and transcription may be interfered by HHP as a result of
disturbance of membrane fluidity. Some researchers reported that high pressure also brings about
the extreme condensation of genetic material in Gram positive bacteria such as L. monocytogenes
and Lactobacillus plantarum as well as Gram negative bacteria such as Salmonella Typhimurium
(Mackey and Manas, 2008). Although HHP does not have any effect on covalent bonds, it has
also been proposed that under high pressure, DNA is more readily cleaved by endonucleases
(Yaldagard et al., 2008).
Intrinsic and extrinsic factors affecting the efficacy of HHP for microbial inactivation
High-pressure inactivation varies as a function of various intrinsic (physiological and
environmental) and extrinsic (processing-related) factors. Physiological factors affecting the
baroresistance of foodborne microbes include the inherent piezoresistance of the organism,
which varies between species and strains as well as its physiological state (age of culture/growth
phase) during exposure to the treatment conditions. Among the various environmental factors
most extensively studied are the growth temperature and physicochemical characteristics of the
medium in which microorganisms are suspended. Since HHP is a multidimensional process, the
extent of microbial inactivation also depends on several processing variables such as exposure
time, temperature and pressure level applied. Intrinsic and extrinsic factors that can affect the

49
response of microorganisms, especially pathogens, to pressure must be considered so that
treatments can be optimized and microbiological safety can be ensured.
Physiological factors affecting microbial pressure sensitivity
Species and strain variation The barosensitivity of different organisms is highly variable. Bacterial endospores,
especially those of Clostridium botulinum, are believed to be the most pressure-resistant life
forms (Gould , 1995). For vegetative forms of bacteria, there is a somewhat feeble correlation
between the Gram stain type/cell morphology and barosensitivity. Gram-positive and Gram-
negative bacteria differ significantly in terms of the chemical structure of their cell walls. In
general, the higher the complexity of an organism, the greater the sensitivity to pressure
treatment. With more complex cell envelopes and weaker cell walls, Gram-negative bacteria are
generally thought to be more pressure-sensitive than Gram-positive bacteria. Chen (2007)
compared the pressure-resistance of various foodborne pathogens suspended in milk and did not
observe any clear trend in pressure resistance between Gram-negative and Gram-positive
organisms. The author reported the order of strains tested, from most to least pressure sensitive
being V. parahaemolyticus > Yersinia enterocolitica > L. monocytogenes > S. Typhimurium >
Salmonella Enteritidis > E. coli O157:H7 > S. aureus > Shigella flexneri. For a given species or
serovar, different strains sometimes have very different sensitivity to high pressure. Frustoli et al.
(2003) inoculated Listeria monocytogenes into an artificial food system and observed differences
among strains, pointing out that the behavior of given strains should not be generalized to other
strains of the same species.

50
Age of cells Bacterial growth phase also has a bearing on pressure sensitivity. Generally speaking, cells
in the exponential phase are more susceptible to pressure than those in the stationary phase of
growth (Palou et al., 1999). Pagan and Mackey (2000) observed that loss of viability was
correlated with a permanent loss of membrane integrity in logarithmic-growth phase cells,
whereas injured membranes of stationary-phase cells were able to repair following pressure
treatment. The authors observed that cell envelopes of exponential phase cells underwent
permanent deleterious changes unseen in stationary-phase cells. Moreover, stationary phase cells
tend to synthesize an array of proteins that protect against various stresses such as elevated
salinity, high temperatures and oxidative damage (Benito et al., 1999). Several authors also
attributed the higher piezotolerance of bacterial cells to the activation of stationary-phase
functions such as the RpoS and SigB, involved in regulating general bacterial stress responses in
Gram negative and Gram-positive cells, respectively (Mackey and Manas, 2008).
Environmental (product-related) factors influencing microbial sensitivity to high pressure
Food composition
Literature has revealed that the high-pressure resistance of microorganisms increases as a
function of the substrate complexity. Generally speaking, broths and real food systems tend to
confer baroprotection to a greater extent than simpler systems such as buffer (Patterson, 2005).
Differential pressure resistance in E. coli O157:H7 and L. monocytogenes were observed in UHT
milk compared to buffer (Patterson, 2005). It is thought that milk is particularly protective for
bacteria such as E. coli and Salmonella, due to the presence of divalent ions as a

51
result of its high calcium content (Garcia-Graells et al., 1999) as well as the colloidal effect of
calcium phosphate (Hauben et al., 1997). Foods other than milk such as liquid egg, oysters and
meats are also generally piezoprotective (Smiddy et al., 2005). The specific baroprotective role
of individual representative food components such as bovine serum albumin (protein), glucose
(carbohydrate) and olive oil (lipid) was also shown to significantly impact on the pressure
resistance of L. monocytogenes (Patterson, 2005). Taken together, it is believed that the
composition of food systems, the combination of specific piezoprotective food components
(protein, carbohydrate and lipid content) and the general physicochemical characteristics of
foods can severely impact on the baroresistance of suspended bacterial cells.
pH
The pH of the food is one of the main factors affecting the growth and survival of
microorganisms; all microorganisms have a pH range in which they can grow and an optimum
pH at which they can grow best. The pH of a food, if not optimal for a particular species, can
thus not only enhance inactivation during treatment but also inhibit outgrowth of sub-lethally
injured cells (Hogan et al., 2005).
Extent of pressure-induced inactivation will generally be enhanced and recovery of sub-
lethally injured cells inhibited for most species, at acidic values. For example the pressure
resistance of E. coli O157:H7 in orange juice is dependent on the pH of the juice, the degree of
inactivation increasing as pH increases; survival of E. coli O157:H7 in orange juice during
storage is also dependent on pH (Linton and Patterson, 2000). Bacterial spores are generally
more resistant to pressure, however HHP in combination with low pH can be sporicidal (Smelt,

52
1998). Compression of foods during HHP treatment may also shift the pH of food as a function
of applied pressure; the direction of pH shift and its magnitude varies depends on the treatment
process under consideration. HHP application to water, weak acid and several buffers is known
to induce a pH decrease by 0.2-0.5 pH units/100 MPa. pH shift during compression can
effectively narrow the pH range for optimal growth or survival of the microorganisms and
potentially enhance pressure inactivation of microorganisms (Marquis, 1976). Ideally, real food
systems should be used as the menstruum to avoid overestimation of microbial inactivation.
Water activity (aw)
Reducing the aw protects microbes against inactivation by HHP. Hence, the water activity
of the product of interest significantly impacts on the bacterial pressure-sensitivity. Oxen and
Knorr (1993) previously showed that an increase in the water activity from 0.94-0.96 to 0.98-1.0
brought about a concomitant increase in the microbial inactivation ratios. The authors observed
that HHP treatment at 400 MPa for 15 minutes at room temperature inactivated the yeast
Rhodotorula rubra when the aw of the suspension was higher than 0.96, whereas the number of
survivors was higher when the aw was depressed (Oxen and Knorr, 1993). The increased
baroresistance of microorganisms at low aw may also be attributed to partial cell dehydration due
to the osmotic pressure gradients between the internal and external fluids, causing cells to shrink,
acquiring thicker membranes, and an increased pressure resistance.
Extrinsic (process-related) factors influencing microbial sensitivity to high pressure
Pressure level
The level of applied pressure is an important factor that not only affects the extent of

53
microbial and enzyme inactivation but also ultimately affects the quality of the treated product.
Generally, increasing the pressure magnitude will generally increase the number of organisms
inactivated. Although pressure levels as high as 1000 MPa have been used in food processing
(Olsson, 1995), the typical pressure range of high-pressure equipment for food processing is
around 600 MPa (Bermudez-Aguirre and Barbosa-Canovas, 2011). Lower pressure may be more
beneficial in terms of destruction of microbial spores than higher pressure because they may
induce spores to germinate that can be more readily inactivated by HHP.
Pressurization mode
HHP can either be applied continuously/statically (pressure hold mode) or dynamically
(cyclic mode). The former implies that the pressure level is increased to a desired level followed
by holding for a specified length of time prior to depressurization. The primary advantage of the
static mode of pressure processing is that once the required pressure level is attained, no further
input energy is required. The parameters that will affect its efficacy are the pressure level and the
pressure holding duration. Cyclic pressurization refers to successive series of pressurization and
depressurization applied at a desired pressure level for a defined holding time. Similar to static
pressurization, the lethal effect also varies as a function of the pressure level. The lethal effect of
pulsed pressurization on microorganisms is thought to be due to the sudden release of high
pressure resulting in lethal pressure shock or bursting of cell walls that kill the microbes (Mussa,
1999). For bacterial spores, effective pulse pressurization has been observed at elevated
temperatures of about 70°C which was thought to soften the cell coat (Mussa, 1999). It was also
postulated that a quick decompression of pressurized microbial cells will cause an adiabatic

54
expansion of water thus resulting in a lethal impact (Munoz et al., 2007).
Treatment holding time
A HHP treatment time include the come-up time (period necessary to reach treatment
pressure), holding time and pressure-release time. Long come-up times will add appreciably to
the total process time and thus affect the product throughput (Hogan et al., 2005). However, at
the same time, the come-up and come-down periods will also contribute to the overall lethality of
the process. Shorter treatment times are generally preferred to make high pressure processing
economically feasible, hence commercial treatment times typically range from seconds to 20
minutes. One way of shortening the treatment time is to apply a higher pressure level although
this may entail a higher operating cost. In all cases, consistency and control of treatment times
are of vital importance in HHP process development (Mussa, 1999).
Temperature
The temperature during pressurization can have an appreciable effect on microbial
inactivation. Several authors have observed that the pressure tolerance by vegetative
microorganisms is highest at normal temperatures (15-30°C). While many HHP processes are
performed at ambient temperature, increasing the treatment temperature (> 30°C) will generally
increase the effectiveness of the HHP treatment (Palou et al., 1999). Ogawa et al. (1990)
demonstrated an increase in the inactivation ratio of naturally present as well as artificially
contaminated microorganisms in mandarin juice treated at 40°C at pressures ranging from 400-
450 MPa. Cheftel (1995) and Carlez et al. (1994) mentioned that temperature-enhanced pressure
inactivation can help lower the minimum pressure level and the treatment time required, which

55
are two major cost determinants. The impact of treatment temperature seems to be related to the
effect of temperature on membrane fluidity as well as protein stability (Carlez et al., 1994). The
documented effect of low treatment temperatures (< 20°C) on pressure inactivation of microbes
is more variable. While the effect of elevated temperatures on pressure inactivation is widely
acknowledged as stated previously, the effect of HHP at reduced temperatures has been shown to
be inconsistent across studies. Carlez et al. (1994) tested the efficacy of high pressure
inactivation (50-400 MPa) of Citrobacter freundii, Pseudomonas fluorescens and Listeria
innocua in inoculated minced beef muscle for 20 min at 4, 20, 35 and 50°C. The author also
observed that the psychrotrophic bacteria (P. fluorescens and L. innocua) were more sensitive to
the effects of pressure at low temperatures compared to ambient temperatures, while
thermotolerant C. freundii was more sensitive at 35 and 50°C. Taken together, these findings
indicate that the influence of low treatment temperature on pressure inactivation is likely to vary
considerably between different microorganisms.
Summary of literature review
Alfalfa sprouts, which are widely consumed in salads and sandwiches, are considered a
serious food safety problem in the US. There have been numerous foodborne disease outbreaks
due to consumption of microbially contaminated sprouts in recent years. The contamination is
mostly carried over from the seeds to the sprouts. Additionally, contamination of irrigation water
used during sprouting could be a source of initial contamination or a vehicle for subsequent
cross-contamination. Decontamination treatments involving non-thermal methods such as high
pressure-processing may offer alternative methods for reduction of pathogenic load on seeds and

56
sprouts. Recent studies on the efficacy of high pressure processing to decontaminate seeds was
reported with variable success depending on the type of sprouting seeds and the target organism.
The efficacy of HHP as an intervention thus warrants more in-depth investigation to quantify its
lethal impact on pathogenic microorganisms that pose greatest concern for sprouts and at the
same time identify the factors that influence the viability and vigor of seeds. Because it is felt
that disinfectants are not very effective in killing pathogens because of their reduced
penetrability into cracks and crevices of seeds/sprouts, it is essential to develop new and highly
effective methods of disinfection.
REFERENCES Andrews, W.H., Mislivec, P.B., Wilson, C.R., Bruce, V.R., Poelma, P.L., Gibson, R., Trucksess, M.W., Young, K., 1982. Microbial hazards associated with bean sprouting. Journal of the Association of Official Analytical Chemists 65, 241-248. Ariefdjohan, M.W., Nelson, P.E., Singh, R.K., Bhunia, A.K., Balasubramaniam, V.M., Singh, N., 2004. Efficacy of high hydrostatic pressure treatment in reducing Escherichia coli O157:H7 and Listeria monocytogenes in alfalfa seeds. Journal of Food Science 69, 117-126. Barak, J.D., Whitehand, L.C., Charkowski, A.O., 2002. Differences in attachment of Salmonella enterica serovars and Escherichia coli O157: H7 to alfalfa sprouts. Applied and Environmental Microbiology 68, 4758-4763. Bari, M.L., Nazuka, E., Sabina, Y., Todoriki, S., Isshiki, K., 2003. Chemical and irradiation treatments for killing Escherichia coli O157:H7 on alfalfa, radish, and mung bean seeds. Journal of Food Protection 66, 767-774. Bartlett, D.H., 2002. Pressure effects on in vivo microbial processes. Biochimica et Biophysica Acta 1595, 367–381. Bass, L.N., Gunn, C.R., Hesterman, O.B., Roos, E.E., 1988. Seed physiology, seedling performance and seed sprouting. Agronomy Monograph 29, 961-983. Beales, N., 2004. Review of the microbiological risks associated with sprouted seeds. Campden and Chorley Food Research Association Group (CCFRA), Review No. 41.

57
Bermudez-Aguirre, D.A., Barbosa-Canovas, G., 2011. An update in High Hydrostatic Pressure, from the Laboratory to Industrial applications. Food Engineering Reviews 3, 44-61. Beuchat, L.R., 1996. Pathogenic microorganisms associated with fresh produce. Journal of Food Protection 59, 204-216. Benito, A., Ventoura, G., Casadei, M., Robinson, T., Mackey, B., 1999. Variation in resistance of natural isolates of Escherichia coli O157:H7 to high hydrostatic pressure, mild heat, and other stresses. Applied and Environmental Microbiology 65, 1564-1569. Burland, V., Shao, Y., Perna, N.T., Plunkett, G., Sofia, H.J., Blattner, F.R., 1998. The complete DNA sequence and analysis of the large virulence plasmid of Escherichia coli O157:H7. Nucleic Acids Research 26, 4196 - 4204. Cai, Y., Ng, L.K., Farber, J.M., 1997. Isolation and characterization of nisin-producing Lactococcus lactis subsp. lactis from bean sprouts. Journal of Applied Microbiology 83, 499-507. Callister, S.M., Agger, W.A., 1987. Enumeration and characterization of Aeromonas hydrophila and Aeromonas caviae isolated from grocery store produce. Applied and Environmental Microbiology 53, 249-253. Carlez, A., Rosec, J.P., Richard, N., Cheftel, J.C., 1994. Bacterial growth during chilled storage of pressure treated minced meat. Lebensmittel Wissenschaft und Technologie 27, 48-54. Castro-Rosas, J., Escartin, E.F., 2000. Survival and growth of Vibrio cholerae O1, Salmonella typhi, and Escherichia coli O157:H7 in alfalfa sprouts. Journal of Food Science 65, 162-165. Castro-Rosas, J., Escartin, E.F., 1999. Incidence and germicide sensitivity of Salmonella Typhi and Vibrio cholerae O1 in alfalfa sprouts. Journal of Food Safety 19, 137-146. Chapman, 2011. Food Safety Network. http://www.foodsafety.ksu.edu/en/article-details.php?a=3&c=14&sc=119&id=865 Chapman, B., 2009. Sprouts associated outbreaks in North America, 1990-2009. Accessed on from Ben Chapman Bites, Food Safety Network. http://foodsafety.k-state.edu/en/article-details.php?a_2&c_6&sc_36&id_865. Charkowski, A.O., Sarreal, C.Z., Mandrell, R.E., 2001. Wrinkled alfalfa seeds harbor more aerobic bacteria and are more difficult to sanitize than smooth seeds. Journal of Food Protection 64, 1292-1298.

58
Cheftel, J.C., Culioli, J., 1997. Effects of high pressure on meat: a review. Meat Science 46, 211-236. Chen, H., 2007. Temperature-assisted pressure inactivation of Listeria monocytogenes in turkey breast meat. International Journal of Food Microbiology 117, 55-60. Coburn, F.D., 1907. The book of alfalfa: history, cultivation and merits. Its uses as a forage and fertilizer. Orange Judd Company. D'Aoust, J.Y., 2001. Foodborne salmonellosis: current international concerns. Food Safety Magazine 7, 10. De Roever, C., 1998. Microbiological study evaluations and recommendations on fresh produce. Food Control 9, 321-347. Delaquis, P.J., Sholberg, P.L., Stanich, K., 1999. Disinfection of mung bean seed with gaseous acetic acid. Journal of Food Protection 62, 953-957. Dong, Y., Iniguez, A.L., Ahmer, B.M.M., Triplett, E.W., 2003. Kinetics and strain specificity of rhizosphere and endophytic colonization by enteric bacteria on seedlings of Medicago sativa and Medicago truncatula. Applied and Environmental Microbiology 69, 1783-1790. Doyle, M.P., 1991. Escherichia coli O157:H7 and its significance in foods. International Journal of Food Microbiology 12, 289-302. Enomoto, K., Takizawa, T., Ishikawa, N., Suzuki, T., 2002. Hot-water treatments for disinfecting alfalfa seeds inoculated with Escherichia coli ATCC 25922. Food Science and Technology Research 8, 247-251. Fan, X., Thayer, D., Sokorai, K.J.B., 2004. Changes in growth and antioxidant status of alfalfa sprouts during sprouting as affected by gamma irradiation of seeds. Journal of Food Protection 67, 561. Fellows, P., 2000. Food processing technology: principles and practice. CRC Press, Cambridge, England. Fett, W.F., 2005. Interventions to ensure the microbiological safety of sprouts. In: Sapers, G.M., Gorny, J.R., Yousef, A.E. (Eds.), Microbiology of Fruits and Vegetables, CRC Press, Boca Raton, FL., p. 187-209. Fett, W.F., Fu, T.J., Tortorello, M.L., 2006. Seed sprouts: the state of microbiological safety. In:

59
Matthews, K.R. (Ed.), Microbiology of Fresh Produce. ASM Press, American Society for Microbiology, 1752 N. St. NW, Washington, D.C. 20036-2904. Food and Drug Administration, 1999a. Guidance for industry: reducing microbial food safety hazards for sprouted seeds and guidance for industry: sampling and microbial testing of spent irrigation during sprout production. Federal Registry 64, 57893-57902. Food and Drug Administration, 1999b. Guidance for Industry: Sampling and Microbial Testing of Spent Irrigation Water During Sprout Production. http://www.cfsan.fda.gov/~dms/sprougd2.html. Food and Drug Administration, 2000. Irradiation in the production, processing and handling of food. Federal Registry 65, 64605-64607. Food and Drug Administration, 2004. Produce safety from production to consumption: Action Plan to minimize foodborne illness associated with fresh produce consumption. Food and Drug Administration. http://www.cfsan.fda.gov/~dms/fc01-1.html Franz, E., Visser, A. A., Diepeningen, A.D., Klerks, M.M., Termorshuizen, A.J., Bruggen, A.H.C., 2007. Quantification of contamination of lettuce by GFP-expressing Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium. Food Microbiology 24, 106-112. Frustoli, M., Gola, S., Miglioli, L., Rovere, P., 2003. Combined heat/high pressure treatments of Listeria spp. in a model system. Industria Conserve 78, 169-182. Garcia-Graells, G.C., Masschalck, B., Michiels, C.W., 1999. Inactivation of Escherichia coli in milk by high hydrostatic pressure treatment in combination with antimicrobial peptides. Journal of Food Protection 62, 1248– 1254. Gould, G.W., 1995. The microbe as a high-pressure target. In: Ledward, D.A., Johnston, D.E., Earnshaw, R.G., Hasting, A.P.M., (Eds.), High Pressure Processing of Foods, Loughborough, Nottingham University Press, p. 27-36. Hallman, J., Quadt-Hallman, A., Mahaffee, W.F., Kloepper. J. W., 1997. Bacterial endophytes in agricultural crops. Canadian Journal of Microbiology 43, 895-914. Hansen-Wester, I., Hensel, M., 2001. Salmonella pathogenicity islands encoding type III secretion systems. Microbes and Infection 3, 549-559. Hara-Kudo, Y., Konuma, H., Iwaki, M., Kasuga, F., Sugita-Konishi, Y., Ito, Y., Kumagai, S., 1997. Potential hazard of radish sprouts as a vehicle of Escherichia coli O157:H7. Journal of Food Protection 60, 1125-1127.

60
Hauben, K.J.A., Bartlett, D.H., Soontjens, C.C.F., Cornelis, K., Wuytack, E.Y., Michiels, C.W., 1997. Escherichia coli mutants resistant to inactivation by high hydrostatic pressure. Applied and Environmental Microbiology 63, 945-950. Heremans, K. 1995. High pressure effects on biomolecules. In: Ledward, D.A., Johnston, D.E., Earnshaw, R.G., Hasting, A.P.M. (Eds.), High Pressure Processing of Foods, Loughborough, Nottingham University Press, p. 81-98. Hite, B.H., 1899. The effect of pressure in the preservation of milk: a preliminary report, Bulletin / West Virginia Agricultural Experiment Station; 58. Morgantown, W.V.: West Virginia Agricultural Experiment Station, p. 15. Hogan, E., Kelly, A., Sun, D.W., 2005. High pressure processing of foods: an overview. In: Sun, D.W. (Ed.), Emerging technologies for food processing, Dublin, Ireland, p. 3–32. Holt, J., Krieg, N., Sneath, P., Staley, J., Williams, S., 1994. Group 5, facultatively anaerobic gram-negative rods. In: Bergey’s Manual of Determinative Bacteriology. Williams & Wilkins (Eds.), Baltimore, MD, USA, p. 175-201. Hoover, D.G., 1993. Pressure effects on biological systems. Food Technology 47, 150-155. Howard, M.B., Hutcheson, S.W., 2003. Growth dynamics of Salmonella enterica strains on alfalfa sprouts and in waste seed irrigation water. Applied and Environmental Microbiology 69, 548-553. Institute of Food Technologists, 2002. Emerging microbiological food safety issues: implications for control in the 21st century, IFT expert report. Chicago, IL. International Food Safety Network, 2001. http://www.foodsafety.ksu.edu/en/article-details.php?%20a=2&c=6&sc=36&id=865 International Specialty Supply, 2011. http://www.sproutnet.com/sprouting_seed_varieties.htm International Sprout Growers Association, 2004. http://www.fda.gov/ohrms/DOCKETS/dailys/04/july04/072804/04n-0258-c00009-vol1.pdf Itoh, Y., Sugita-Konishi, Y., Kasuga, F., Iwaki, M., Hara-Kudo, Y., Saito, N., Noguchi, Y., Konuma, H., Kumagai, S., 1998. Enterohemorrhagic Escherichia coli O157:H7 present in radish sprouts. Applied and Environmental Microbiology 64, 1532-1535. Jaquette, C.B., Beuchat, L.R., Mahon, B.E., 1996. Efficacy of chlorine and heat treatment in killing Salmonella Stanley inoculated onto alfalfa seeds and growth and survival of the pathogen

61
during sprouting and storage. Applied and Environmental Microbiology 62, 2212-2215. Kadharmeston, C., 1998. Thermal property and functionality of whey protein concentrate treated by heat or high hydrostatic pressure. Master Thesis. Washington State University, USA. Knorr, D., 1993. Effects of high-hydrostatic pressure processes on food safety and quality. Food Technology 47, 156. Kodama, T., Akeda, Y., Kono, G., Takahashi, A., Imura, K., Lida, T., Honda, T., 2002. Cellular Microbiology 4, 213-222. Kothary, M.H., Babu, U.S., 2001. Infective dose of food-borne pathogens in volunteers: a review. Journal of Food Safety 21, 49-73. Knutton, S., Baldwin, T., Williams, P.H., McNeish, A.S., 1989. Actin accumulation at sites of bacterial adhesion to tissue culture cells: basis of a new diagnostic test for enteropathogenic and enterohemorrhagic Escherichia coli. Infections and Immunity 57, 1290-1298. Linton M., Patterson, M.F., 2000. High pressure processing of foods for microbiological safety and quality. Acta Microbiologica et Immunologica Hungarica 47, 175-182. Mackey, B.M., Manas, P., 2008. Inactivation of Escherichia coli by high pressure. In: Michiels, C.W., Bartlett, D.H., Aertsen, A., (Eds.), High-Pressure Microbiology, ASM Press. Washington, DC, p. 53-85. Mahon, B.E., Ponka, A., Hall, W., Komatsu, K., Beuchat, L., Shiflett, S., Siitonen, A., Cage, G., Lambert, M., Hayes, P., Bean, N., Griffin, P., Slutsker, L., 1997. An international outbreak of Salmonella infections caused by alfalfa sprouts grown from contaminated seeds. Journal of Infectious Diseases 175, 876-882. Manãs, P., Mackey, B. M., 2004. Morphological and physiological changes induced by high hydrostatic pressure in exponential and stationary phase cells of Escherichia coli: relationship with cell death. Applied and Environmental Microbiology 70, 1545-1554 Marler, 2011. Food Poison Journal. http://www.foodpoisonjournal.com/foodborne-illness-outbreaks/was-world-wide-e-coli-o104h4-outbreak-caused-by-sprouts/ Marquis, R.E., 1976. High-pressure microbial physiology. In: Rose, A.H., Tempest, D.W., (Eds.), Advances in Microbial Physiology, Academic Press, New York, p. 105-108. Matos, A., Garland, J.L., 2005. Effects of community versus single strain inoculants on the biocontrol of Salmonella and microbial community dynamics in alfalfa sprouts. Journal of Food Protection 68, 40-48.

62
McDaniel, T.K., Jarvis, K.G., Donnenberg, M.S., Kaper, J.B., 1995. A genetic locus of enterocyte effacement conserved among diverse enterobacterial pathogens. Proceedings of the National Academy of Sciences USA 92, 1664-1668 McMahon, M.A.S., Wilson, I.G., 2001. The occurrence of enteric pathogens and Aeromonas species in organic vegetables. International Journal of Food Microbiology 70, 155-162. Mertens, B., 1995. New Methods of Food Preservation, Blackie Academic and Professional, New York, p. 135. Meyer, D., 1999. Alfalfa: Seed germination seedling and growth vegetative development. North Dakota State University, Agricultural Experiment Station. Meyerowitz, S., 2007. Sprouts the miracle food: the complete guide to sprouting. 6th ed. Sproutman Publications, PO Box 1100, Great Barrington, Mass. 01230. Meyerowitz, S., 1999. Sprouts: the miracle food. Sproutman Publications, Massachusetts, U.S.A., pp. 57-79. Molbak, K., Olsen, J.E., Wegener, H.C., 2006. Salmonella infections. In: Riemann, H.P., Cliver, D.O. (Eds.), Foodborne infections and intoxications, Academic Press, Elsevier, San Diego, CA, p. 57-115. Munoz, M., Ancos, B.D., Sanchez-Moreno, C., Cano, M.P., 2007. Effects of High Pressure and mild heat on endogenous microflora and on the inactivation and sublethal injury of Escherichia coli inoculated into fruit juices and vegetable soup. Journal of Food Protection 70, 1587-1593. Mussa, D.M., 1999. High pressure processing of milk and muscle foods: evaluation of process kinetics, safety and quality changes. PhD Dissertation, McGill University, Montreal, Canada. National Advisory Committee on Microbiological Criteria for Foods (NACMCF), 1999. Microbiological safety evaluations and recommendations on sprouted seeds. International Journal of Food Microbiology 14, 185-246. Olsson, S., 1995. Production equipment for commercial use. In: Ledward D.A., Johnston, D.E., Earnshaw, R.G., Hasting, A.P.M., (Eds.), High Pressure Processing of Foods, Nottingham University Press, Nottingham, p. 167. Otero, L., Molina-Garcia, A.D., Sanz, P.D., 2000. Thermal effect in foods during quasi-adiabatic pressure treatments. Innovative Food Science and Emerging Technologies 1, 119-126.

63
Oxen, P., Knorr, D., 1993. Baroprotective effects of high solute concentrations against inactivation of Rhodotorula rubra. Lebensmittel Wissenschaft und Technologie 26, 220-223. Pagan, R., Mackey, B., 2000. Relationship between membrane damage and cell death in pressure-treated Escherichia coli cells: differences between exponential- and stationary-phase cells and variation among strains. Applied and Environmental Microbiology 66, 2829-2834. Palmai, M., Buchanan, R.L., 2002. Growth of Listeria monocytogenes during germination of alfalfa sprouts. Food Microbiology 19, 195-200. Palou, E., Lopez-Malo, A., Barbosa-Canovas, G.V., Swanson, B.G., 1999. High pressure treatment in food preservation. In: Rahman, M.S. (Ed.), Handbook of food preservation, Marcel Dekker, New York, NY, pp. 533-576. Pao, S., Kalantari, A., Khalid, M.F., 2008. Eliminating Salmonella enterica in alfalfa and mung bean sprouts by organic acid and hot water immersions. Journal of Food Processing and Preservation. 32, 335-342. Park, S.W., Sohn, K.H., Shin, J.H., Lee, H.J., 2001. High hydrostatic pressure inactivation of Lactobacillus viridescens and its effects on the ultrastructure of cells. International Journal of Food Science and Technology 36, 775-781. Paton, J.C., Paton, A.W., 1998. Pathogenesis and diagnosis of Shiga Toxin-producing Escherichia coli infections. Clinical Microbiology Reviews 11, 450-479. Patterson, M.F., Linton, M., Doona, C.J., 2007. Introduction to high pressure processing of foods. In Doona, C.J., Dunne, C.P., Feehery, F.E., (Eds.), High Pressure Processing of Foods, Iowa, Blackwells Publishing, USA, p. 1-14. Patterson, M.F., 2005. A review: microbiology of pressure treated foods. Journal of Applied Microbiology 98, 1400-1409. Patterson, J.E., Woodburn, M.J., 1981. Klebsiella and other bacteria on alfalfa and bean sprouts at the retail level. Journal of Food Science 45, 492-495. Peñas, E., Gomez, R., Frias, J., Vidal-Valverde, C., 2008. Application of high-pressure treatment on alfalfa (Medicago sativa) and mung bean (Vigna radiata) seeds to enhance the microbiological safety of their sprouts. Food Control 19, 698-705. Piernas, V., Guiraud, J.P., 1998. Control of microbial growth on rice sprouts. International Journal of Food Science and Technology 33, 297-305.

64
Pierre, P.M-M., 2004. Efficacy of chemical sanitizers to inactivate Escherichia coli O157:H7, Salmonella Typhimurium DT 104 and Listeria monocytogenes on alfalfa seeds and sprouts. Masters Thesis, Michigan State University, USA. Portnoy, B.L., Goepfert, J.M., Harmon, S.M.,1976. Outbreak of Bacillus cereus Food Poisoning Resulting from Contaminated Vegetable Sprouts. American Journal of Epidemiology 103, 589-594. Powell, D., Blaine, K., Chapman, B., 2002. Sprouts in Canada. Commentary from the Food Safety Network. http://www.sproutnet.com/Press/sprouts_in_canada.htm Price, T.V., 1988. Seed sprout production for human consumption - a review. Canadian Institute of Food Science and Technology Journal 21, 57-65. Prokopowich, D., Blank, G., 1991. Microbiological evaluation of vegetable sprouts and seeds. Journal of Food Protection 54, 560-562. Puohiniemi, R., Heiskanen, T., Siitonen, A., 1997. Molecular epidemiology of two international sprout-borne Salmonella outbreaks. Journal of Clinical Microbiology 35, 2487-2491. Rajkowski, K.T., Thayer, D.W., 2001. Alfalfa seed germination and yield ratio and alfalfa sprout microbial keeping quality following irradiation of seeds and sprouts. Journal of Food Protection 64, 1988-1995. Rastogi, N.K., Raghavarao, K.S.M.S., Balasubramaniam, V.M., Niranjan, K., Knorr, D., 2007. Opportunities and challenges in high pressure processing of foods. Critical Reviews in Food Science and Nutrition 47, 69–112. Robertson, L.J., Johannessen, G.S., Gjerde, B.K., Loncarevic, S., 2002. Microbiological analysis of seed sprouts in Norway. International Journal of Food Microbiology 75, 119-126. Scallan, E., Hoekstra, R.M., Angulo, F.J., Tauxe, R.V., Widdowson, M-A, Roy, S.L., Jones, J.L., Griffin, P.M., 2011. Emerging Infectious Diseases 17, 7-15 Scouten, A.J., Beuchat, L.R., 2002. Combined effects of chemical, heat and ultrasound treatments to kill Salmonella and Escherichia coli O157:H7 on alfalfa seeds. Journal of Applied Microbiology 92, 668-674. Sellman, P., Sellman, G., 1981. The complete sprouting book. Turnstone Press Limited. Sharma, R.R., Fett, W.F., Beuchat, L.R., Demirci, A., 2003. Application of ozone for inactivation of Escherichia coli O157:H7 on inoculated alfalfa sprouts. Journal of Food

65
Processing and Preservation 27, 51-64. Sharma, R., 2002. Decontamination of alfalfa seeds and sprouts using novel methods. PhD Dissertation, Penn State University, USA. Sivapalasingam, S., Friedman, C.R., Cohen, L., Tauxe, R.V., 2004. Fresh produce: a growing cause of outbreaks of foodborne illness in the United States, 1973 through 1997. Journal of Food Protection 67, 2342-2353. Smelt, J.P.P.M., 1998. Recent advances in the microbiology of high pressure processing. Trends in Food Science and Technology 9, 152-158. Smiddy, M., O’Gorman, L., Sleator, D.R, Kerry, P.J., Patterson, F.M., Kelly, L.A., Hill, C., 2005. Greater high-pressure resistance of bacteria in oysters than in buffer. Innovative Food Science and Emerging Technologies 6, 83–90. Sperandio, V., Kaper, J.B., Bortolini, M.R., Neves B.C., Keller, R., Trabulsi, L.R., 2006. Charcaterization of the locus of enterocyte effacement (LEE) in different enteropathogenic Escherichia coli (EPEC) and Shiga-toxin producing Escherichia coli (STEC) serotypes. FEMS Microbiology Letters 164, 133-139. Stewart, D., Reineke, K., Ulaszek, J., Fu, T., Tortorello, M., 2001. Growth of Escherichia coli O157:H7 during sprouting of alfalfa seeds. Letters in Applied Microbiology 33, 95-99. Taormina, P.J., Beuchat, L.R., 1999. Behavior of enterohemorrhagic Escherichia coli O157:H7 on alfalfa sprouts during the sprouting process as influenced by treatments with various chemicals. Journal of Food Protection 62, 850-856. Taormina, P.J., Beuchat, L.R., Slutsker, L., 1999. Infections associated with eating seed sprouts: An international concern. Emerging Infectious Diseases 5, 626-634. Tauxe, R.V., 1997. Emerging foodborne diseases: an evolving public health challenge. Dairy, food and environmental sanitation : a publication of the International Association of Milk, Food and Environmental Sanitarians 17, 788-795. Tauxe, R.V., 1991. Salmonella - a postmodern pathogen. Journal of Food Protection 54, 563-568. Thayer, D.W., Rajkowski, K.T., Boyd, G., Cooke, P.H., Soroka, D.S., 2003. Inactivation of Escherichia coli O157:H7 and Salmonella by gamma irradiation of alfalfa seed intended for production of food sprouts. Journal of Food Protection 66, 175-181.

66
Ting, E.Y., Marshall, R.G., 2002. Production issues related to UHP food. In: Welti-Chanes, J., Barbosa-Canovas, G.V., Aguilera, J.M. (Eds.) Engineering and Food for the 21st century. Food Preservation Technology Series, CRC Press, Boca Raton, Ch. 44, p. 727-738. Tzipori, S., Karch, H., Wachsmuth, I.K., Robins-Browne, R.M., O’Brien, A.D., Lior, H., Cohen, M.L., Smithers, J., Levine, M.M., 1987. Role of a 60-megadalton plasmid and Shiga-like toxins in the pathogenesis of infection caused by enterohemorrhagic Escherichia coli O157:H7 in gnotobiotic pigs. Journal of Infectious Diseasesa 154, 712-716. Wang, 2006. Simultaneous quantitation of Escherichia coli O157:H7, Salmonella and Shigella in ground beef by multiplex real-time PCR and immunomagnetic separation. Masters Thesis submitted to University of Missouri-Columbia. Warriner, K., Spaniolas, S., Dickinson, M., Wright, C., Waites, W.M., 2003. Internalization of bioluminescent Escherichia coli and Salmonella Montevideo in growing bean sprouts. Journal of Applied Microbiology 95, 719-727. Weagant, S.D., Bound, A.J., 2001. Evaluation of techniques for enrichment and isolation of Escherichia coli O157:H7 from artificially contaminated sprouts Weissinger, W.R., Beuchat, L.R., McWatters, K.H., 2001. Evaluation of volatile chemical treatments for lethality to Salmonella on alfalfa seeds and sprouts. Journal of Food Protection 64, 442-450. Wigmore, A. 1986. The Sprouting Book: How to grow and use sprouts to maximize your health and vitality. Avery, a member of Penguin Group (USA) Inc. Wuytack, E.Y., Michiels, C.W., Meersseman, K., Diels, A.M., 2003. Decontamination of seeds for sprout production by high hydrostatic pressure. Journal of Food Protection 66, 918-923. Yaldagard, M., Mortazavi, S.Y., Tabatabaie, F., 2008. The principles of ultra high-pressure technology and its application in food processing/preservation: a review of microbiological and quality aspects. African Journal of Biotechnology 7, 2739-2767.

67
Chapter 3
Potential application of high hydrostatic pressure to eliminate
Escherichia coli O157:H7 on alfalfa sprouted seeds
Published in Int. J. Food Microbiol. 2008. 128, 348-353.

68
Abstract
Sprouts eaten raw are increasingly being perceived as hazardous foods as they have been
implicated in Escherichia coli O157:H7 outbreaks where the seeds were found to be the likely
source of contamination. The objective of our study was to evaluate the potential of using high
hydrostatic pressure (HHP) technology for alfalfa seed decontamination. Alfalfa seeds inoculated
with a cocktail of five strains of E. coli O157:H7 were subjected to pressures of 500 and 600
MPa for 2 min at 20°C in a dry or wet (immersed in water) state. Immersing seeds in water
during pressurization considerably enhanced inactivation of E. coli O157:H7 achieving
reductions of 3.5 log and 5.7 log at 500 and 600 MPa, respectively. When dry seeds were
pressurized, both pressure levels reduced the counts by < 0.7 log. To test the efficacy of HHP to
completely decontaminate seeds whilst meeting the FDA requirement of 5 log reductions, seeds
inoculated with a ~5 log CFU/g of E. coli O157:H7 were pressure-treated at 600 and 650 MPa at
20°C for holding times of 2 to 20 min. A > 5 log reduction in the population was achieved when
600 MPa was applied for durations of ≥ 6 min although survivors were still detected by
enrichment. When the pressure was stepped up to 650 MPa, the threshold time required to
achieve complete elimination was 15 min. Un-inoculated seeds pressure-treated at 650 MPa for
15 min at 20°C successfully sprouted achieving a germination rate identical to untreated seeds
after eight days of sprouting. These results therefore demonstrate the promising application of
HHP on alfalfa seeds to eliminate the risk of E. coli O157:H7 infections associated with
consumption of raw alfalfa sprouts.

69
Introduction
In 1994, the Food and Drug Administration (FDA) and the U.S. Department of
Agriculture (USDA) recommended that Americans increase their consumption of fresh fruits and
vegetables (Hu et al., 2004). Fruit and vegetable consumption has since increased over the past
decade as a result of the heightened awareness of their health benefits, resulting in a significant
growth in the fresh produce industry. However, the incidence of food-borne illness associated
with the consumption of salad vegetables and fruits have also increased. An increase in the
consumption of raw alfalfa sprouts has been paralleled by an upsurge in the number of food-
borne disease outbreaks in recent years (Sharma et al., 2003). Seeds have been attributed to be
the main source of contamination of sprouts, although pathogens may be introduced at various
stages of the spout production continuum (Utlee et al., 1998; NACMCF, 1999; Taormina and
Beuchat, 1999; Ogawa et al., 2000; Weissinger et al., 2001).
Sanitizing sprouted seeds presents a unique challenge in the arena of produce safety in
that even a low residual pathogen population remaining post-treatment appears capable of
growing to very high levels (up to 8 log CFU/g) due to favorable conditions of moisture,
temperature and nutrient availability during seed germination and subsequent sprout growth
(Fett, 2006). In addition, after a sanitizing procedure seed germination as well as spout yield and
quality need to be maintained at commercially acceptable levels. In 1999, based on research
available at that time, the FDA recommended that seeds be disinfested by washing with 20,000
ppm calcium hypochlorite solution prior to sprouting (Rajkowski and Thayer, 2001). This
treatment has been shown to only reach pathogenic microorganisms on the seed surface and there

70
is still no guarantee that all contamination can be removed (Mundt and Hinkle, 1976; Taormina
and Beuchat, 1999). Due to the unreliability of the disinfection step, the FDA requires testing of
spent irrigation water for the pathogens within 24 hours after the start of the sprouting procedure
(U.S. FDA, 1999; Waddell and Troxell, 2000).
Alternative methods thus need to be sought to ensure the safety of sprout consumption
without any detriment to their health-related qualities. High hydrostatic pressure (HHP)
technology could be a useful tool to eliminate pathogenic microorganism like E. coli O157:H7 in
sprouted seeds. HHP acts instantaneously and uniformly throughout the mass of food
independent of the size, shape or geometry (Farkas and Hoover, 2000). Previous research on the
application of HHP to decontaminate alfalfa seeds as carried out by Ariefdjohan (2004) has
shown that pressurizing seeds in the dry state greatly impaired the seed viability. On the other
hand, Penas et al. (2008) soaked alfalfa seeds in water for three hours, decanted the water and
pressure-treated the seeds without water immersion and found a subsequent decrease in the
germination yield with respect to untreated seeds. The overall objective of our study was to
develop a HHP process underscoring the importance of immersing seeds in water during
pressure treatment to completely eliminate a population of 105 CFU/g of E. coli O157:H7
inoculated onto alfalfa seeds while retaining the seeds viability.

71
Materials and Methods
Effect of pressure levels and water-immersion on the germination rate of alfalfa and mung
bean seeds
Alfalfa and mung bean seeds were obtained from International Specialty Supply
(Cookeville, Tenn., USA). One hundred alfalfa or mung bean seeds were placed in individual
plastic pouches (Nylon/Polyethylene, Koch Supplies, Kansas City, MO). Water (2 ml for alfalfa
seeds and 6 ml for mung bean seeds) were added to half of the pouches and those pouches were
double heat-sealed. For seeds in the dry state, seeds were vacuum-packaged. To avoid leakage
during pressure-treatment, each sample pouch was placed in a larger 8-mil thick PVC plastic
pouch (Warp Bros., Chicago, IL, USA). HHP treatment of samples was carried out using a high
pressure unit with temperature control (Model Avure PT-1, Avure Technologies, Kent, WA).
The experiments were conducted at 20°C using water as a hydrostatic medium. Pressurization
was conducted at 200-600 MPa for 2 min. The temperature of the water-bath was monitored
through a K-type thermocouple. The temperature and pressure data were recorded every 2 s
(DASYTEC USA, Bedford, NH). The pressure-come-up rate was approximately 22 MPa/s. The
pressure-release was almost immediate (< 4 s). Pressurization time reported in this study does
not include the pressure come-up or release times. Temperature increase during pressure
treatment due to adiabatic heating was 2.5ºC/100 MPa at 20ºC (Chen et al., 2005). To determine
the germination rate of treated and un-treated seeds, seeds were soaked in DI-water for 3 h and
100 seeds were spread evenly between two pieces of wet paper towels on a plastic rack which in
turn was placed into a water-filled bucket to provide a moist environment for the seeds. The

72
water level was maintained below the seeds’ level. The bucket was covered loosely with a piece
of plastic film to allow exchange of air between the inside and outside of the bucket. The bucket
was kept at room temperature (~21ºC) for 8 days (suggested by the seeds provider). The seeds
were visually evaluated for germination on days 3-8.
Effect of water-immersion of alfalfa seeds on pressure inactivation of E. coli O157:H7
Bacterial Strains
E. coli O157:H7 strain 1730 and Cider strain (Ibrahim et al., 2006), strains 250 and 251
(Bhagwat et al., 2005) (Courtesy of Dr. Kniel, University of Delaware) and J58 (Barak et al,
2005) (Courtesy of Dr Joerger, University of Delaware) were used. The cells of E. coli O157:H7
were adapted to grow in tryptic soy Broth plus 0.6% yeast extract (Difco Laboratories, Sparks,
MD, USA) supplemented with 50 µg/ml of nalidixic acid (Fisher Scientific, Hampton, NH,
USA) (TSBYE-N). Individual cultures were grown in TSBYE-N overnight at 35°C. Cultures
were then transferred (one loopful) into 10 ml of fresh TSBYE-N and incubated at 35°C for 24 h.
Equal volume of individual cultures was mixed to form a five-strain cocktail of E. coli O157:H7.
Preliminary experiments demonstrated that the five-strain wild-type cocktail and the five-strain
nalidixic acid-resistant cocktail of E. coli O157:H7 had comparable pressure sensitivity (data not
shown).
Inoculation of seeds
The cocktail (10 ml) were mixed with 100 ml of sterile 0.1% peptone water (Fisher).
Alfalfa seeds (100 g) was added to the cell suspension and gently stirred for 5 min. The seeds
were separated from the cell suspension by pouring the mixture over a double layer of

73
cheesecloth supported by a wire screen and dried inside a bio-safety hood at room temperature
(21 ± 1°C) for 24 h. Dried seeds with an approximate inoculation level of 109 CFU/g of E. coli
O157:H7 were placed in sterile pouches and stored at 4°C.
Pressure treatment
Inoculated alfalfa seeds (2 g) were bagged and pressure-treated either in the wet or dry
state. For water-immersed seeds, 3 ml of sterile De-ionized (DI) water was added to the seeds.
The seeds were treated at pressures of 500 and 600 MPa for 2 min at 20°C. The counts of E. coli
O157:H7 in pressure-treated dry and wet seed samples and two untreated controls were
determined as described below. The two untreated controls were (i) dry seeds without pressure
treatment and (ii) seeds immersed in 3 ml of water and washed under agitation for 5 min without
pressure treatment.
Microbiological analysis
Pouches containing treated seeds were cut open aseptically. The 2 g seeds were poured
into a stomacher bag to which 8 ml of sterile 0.1% peptone water was added and subsequently
stomached for 2 min at 260 rpm (Seward 400 Stomacher, Seward Medical Co., London, U.K.).
The seed slurry was serially diluted in sterile 0.1% peptone and surface-plated (100 µl) in
duplicate on tryptic soy agar with 0.6% yeast extract (Difco Laboratories, Sparks, MD, USA)
supplemented with nalidixic acid to a final concentration of 50 µg/ml (TSAYE-N). TSAYE-N
plates were incubated at 35ºC for 72 h. Presumptive colonies of E. coli O157:H7 formed on the
plates were enumerated. Occasionally, colonies were confirmed to be E. coli O157:H7 using
either a BAX® system for screening/ E. coli O157:H7 PCR assay (Qualicon-DuPont,

74
Wilmington, DE, USA) or Rapid E. coli O157:H7 Test Methods (Strategic Diagnostics Inc.,
Newark, DE, USA).
Effect of volume of water added to alfalfa seeds on baro-inactivation of E. coli O157:H7
and viability of pressure-treated seeds
Results from the above experiments demonstrated that addition of water to the seeds
during pressure-treatment enhanced both bacterial inactivation and seed germination. Therefore,
the volume of water required during pressure treatment for optimum bacterial inactivation and
simultaneous seed viability retention was determined. Various volumes (1, 2, 3, and 5 ml) of
sterile DI water were added to two grams of inoculated or un-inoculated seeds. The inoculated
and un-inoculated seeds immersed in water were pressure-treated at 600 MPa for 2 min at 20ºC
and subjected to microbiological analysis and germination assay as described above.
Effect of pressure level and treatment time on inactivation of a population of ~105 CFU/g of
E. coli O157:H7 inoculated on alfalfa seeds
In order to show the decontamination efficacy of HHP on seeds that are typically
subjected to a very low level of field-contamination whilst simultaneously demonstrate that the
process could meet the FDA requirement (> 5 log reduction), seeds were inoculated with a lower
inoculation level of ~105 CFU/g. To determine the condition required to achieve complete
lethality, 2 g of inoculated seeds immersed in 3 ml of sterile DI water were subjected to pressure
treatment of 600 and 650 MPa at 20°C for 2-20 min. The 650 MPa was the upper limit possible
for the pressure unit. The counts of E. coli O157: H7 in the treated and untreated seeds were
determined as described above. The remaining seed slurries after plate count determination were

75
enriched in 90 ml of TSBYE-N and incubated for 48 h at 35ºC to allow resuscitation of sub-
lethally injured cells. Samples were streaked onto sorbitol MacConkey agar (Difco Laboratories,
Sparks, MD, U.S.A.) plates supplemented with 50 µg/ml of nalidixic acid. After 24 h incubation,
sorbitol-negative E. coli O157:H7 typified by pale or colorless colonies were identified on the
plates (Hitchins et al., 1995). Colonies were occasionally subjected to confirmation using the
PCR or Rapid Test methods as described above. Since it was found that the treatment of 650
MPa for 15 min at 20°C was able to completely eliminate the pathogen, this treatment was
applied to un-inoculated seeds and the germination rate of the treated and un-treated seeds were
determined.
Statistical analysis
All experiments were replicated at least three times. Where appropriate, statistical
analyses were conducted using Minitab® Release 15 (Minitab Inc., University Park, PA, USA).
One-way analysis of variance (ANOVA) and Tukey’s one-way multiple comparisons were used
to determine differences in the populations of E. coli O157:H7 recovered on treated alfalfa seeds
as well as differences in the germination rate of seeds. Significant differences were considered at
the 95% confidence level (P < 0.05).
Results
Effect of pressure levels and water-immersion on the germination rate of alfalfa and mung
bean seeds
Results are shown in Tables 1 and 2. Mung bean seeds had considerably higher
germination rates when they were immersed in water during pressurization than when they were

76
pressure-treated in the dry state (P < 0.05). The pressure-treated seeds achieved > 85%
germination across all pressure levels after 8 days of germination. The germination rate of
pressure-treated dry mung-bean seeds was severely impacted at all pressure levels and the best
germination yield achieved was 9.7%. For alfalfa seeds, the difference between untreated and
treated wet seeds across all pressure levels became smaller with increased germination time; and
eventually disappeared with no significant difference (P > 0.05) in the germination capacity
between un-treated alfalfa seeds (control) and pressure-treated wet seeds by the eighth day of
germination. The germination rates and final germination percentages of treated dry seeds were
consistently lower than either the control or their immersed counterparts at corresponding
pressure levels. It should be noted that treatment at pressure as high as 600 MPa did not have any
adverse impact on the germination capability and rate of pressure-treated wet seeds.
Effect of water-immersion of alfalfa seeds on pressure inactivation of E. coli O157:H7
Washing the seeds for 5 min in water reduced the counts of E. coli O157:H7 by 0.3 log
CFU/g whilst immersing seeds in water during pressurization considerably enhanced pressure
inactivation of E. coli O157:H7. The pressure treatments reduced the counts of E. coli O157:H7
by 3.5 log and 5.7 log at 500 MPa and 600 MPa, respectively when seeds were immersed in
water during pressurization. When dry seeds were pressurized, both pressure treatments reduced
the counts by ≤ 0.7 log10CFU/g.

77
Effect of volume of water added to alfalfa seeds on baro-inactivation of E. coli O157:H7
and viability of pressure-treated seeds
Since immersion of seeds in water was critical for baro-inactivation of E. coli O157:H7
and viability of seeds, it was logical to treat seeds at 600 MPa immersed in varying volumes of
water to determine the optimum volume required to immerse seeds. The pressure inactivation of
E. coli O157:H7 at different volumes of immersion water are presented in Table 3.3. When
volume of water was increased, there was a commensurate increase in the log reductions of the
pathogen with a maximum of 4.4 log at a water volume of 3 ml. Further addition of water to the
immersed seeds did not enhance pressure inactivation. With respect to seed viability, seeds
pressure-treated whilst immersed in the various volumes of water did not differ significantly (P >
0.05) in their germination percentages (Table 3.3).
Effect of pressure level and treatment time on inactivation of a population of ~105 CFU/g of
E. coli O157:H7 inoculated on alfalfa seeds
The ability of pressure treatment to completely eliminate a population of 105 CFU/g of E.
coli O157:H7 inoculated on alfalfa seeds was investigated. While population reduction through
HHP treatment is desirable, it is more important to achieve complete elimination of the pathogen
since even a few cells can grow explosively to as high as 7-8 log CFU/g in the final sprouts
during germination. Figure 3.1. shows the survival curves for E. coli O157:H7 at 600 and 650
MPa for varying exposure times. There was a close association between bacterial inactivation
and treatment duration; population reductions increased as a function of holding time. A
maximum average reduction of ~5.5 log CFU/g was achieved when the treatment time was ≥ 6

78
min for the 600 MPa treatments. However, none of the treatments at 600 MPa were adequate in
completely eliminating the pathogen as survivors were detected by enrichment. For pressure
treatment at 650 MPa, treatment at ≥ 4 min was able to achieve a 5 log reduction of the pathogen
although survivors were still detected by enrichment when the holding time was ≤ 10 min.
Complete elimination was consistently observed for the 15 min treatment. Un-inoculated seeds
subjected to a treatment of 650 MPa for 15 min at 20°C retained their germination potential with
a final germination yield of 93%, which was identical to the average germination yield of
untreated seeds (Figure 3.2.).
Discussion
Increasing consumer demand for minimally processed, additive-free, shelf-stable
products has prompted food scientists to explore other physical preservation methods as
alternatives to traditional heat treatments. The application of HHP as a physical decontamination
method may be more promising for use on alfalfa seeds since the use of chemical
decontamination is difficult to reconcile with the ‘‘health food’’ image of seed sprouts (Wuytack
et al., 2003). It has been suggested that the bottleneck to most seed decontamination techniques
reside in their inability to reach crevices or cracks where pathogens may be lodged or embryonic
and endospermic tissues where they may become internalized (Beuchat, 1996; NACMCF, 1999).
However, HHP is thought to present unique advantages due to the fact that it acts instantaneously
and uniformly throughout a pressurized sample regardless of size, shape, and geometry (Farkas
and Hoover, 2000). Hence, HHP could act uniformly at all sites within the seeds and on the seed
surface thereby targeting the superficial as well as the internalized pathogens. The application of

79
HHP to decontaminate seeds from pathogenic microorganisms such as Salmonella and E. coli
O157 has been investigated previously (Wuytack et al., 2003; Ariefdjohan et al., 2004; Penas et
al., 2008) with varying degrees of efficacy. Our findings that alfalfa and mung bean seeds
subjected to pressure treatment in the dry state germinated poorly are in agreement with those of
Ariefdjohan et al. (2004). They reported that HHP severely impacted the viability of alfalfa seeds
pressurized in the dry state with a significant correlation between the pressure level used and the
germination rate. They attributed the low germination rate to the inability of the structure of dry
alfalfa seeds to sustain pressures ranging from 275 MPa to 575 MPa. HHP on dry mung bean
seeds was more severe than on dry alfalfa seeds with germination rate varying from 1 to 10%
(Table 3.1 and Table 3.2). Pressure-treated dry alfalfa seeds exhibited minimal visible signs of
damage whilst dry mung bean seeds appeared to have undergone more extensive damage. It is
thought that the mung bean seed tissues have more intercellular air spaces which were
compressed considerably during pressure treatment inducing physical damage to the seeds.
When seeds are immersed in water, they typically retain their overall characteristics better than
when treated in the dry state. Indeed, Michel and Autio (2001) report that pressure acts
instantaneously on a plant tissue at the tissue, cellular and molecular level. A major hurdle for
high pressure treatment of plant tissues is the presence of intercellular air spaces. Because of the
high compressibility of air, the tissue is severely compressed, resulting in cell wall breakage (if
the cell wall is not flexible enough), membrane disruption, loss of compartmentalization, and
liberation of cellular compounds. They recommended circumventing this problem by filling the
intercellular spaces with a liquid of low compressibility such as water (which is only

80
compressible by 15% at 600 MPa) before HHP treatment (Michel and Autio, 2001). When the
seeds were immersed in water, they probably absorbed water from their surrounding as a result
of osmosis and filled up the intercellular air spaces making them better able to withstand the
external hydrostatic pressure.
With regard to pressure inactivation of E. coli O157:H7, the population reductions at 500
and 600 MPa were significantly (P < 0.05) correlated with increasing pressure level when seeds
were pressurized in the immersed state. In addition, the degree of inactivation was significantly
higher when seeds were immersed in water than when they were pressurized in the dry state.
Ariefdjohan et al. (2004) also found that pressure treatment was not effective against E. coli
O157 when inoculated alfalfa seeds were pressure treated in the dry state. A pressure treatment
of 575 MPa for 2 min at approximately 30°C only reduced the population of E. coli O157 in
alfalfa seed by 1.4 log CFU/g. It is thought that water immersion of seeds enhances bacterial
inactivation due to the fact that the pathogens that were previously sequestered in cracks,
crevices or other “microenvironments” of seeds, or trapped beneath the external coat become
exposed at the surface as a result of imbibition of water during pressure treatment. As a result,
these cells which previously benefited from some form of physical protection are now exposed at
the surface or released into the surrounding water rendering them more vulnerable to pressure
inactivation. Moreover, a low water activity is known to confer protection to whole organisms
against heat and pressure (Palou et al., 1999; Smelt et al., 2001). There are numerous reports
showing the protective effects of low water activity against pressure (Oxen and Knorr, 1993;
Palou et al., 1997; Kingsley and Chen, 2008). The lower reductions in the population in dry

81
seeds as opposed to water-immersed seeds at both 500 and 600 MPa may be partially attributed
to the protective effect of the lower water activity in the dry state of seeds.
Since water was found to be a limiting factor affecting both the extent of population
reduction and the viability of seeds, the effect of varying the volume of water immersion during
pressure treatment was investigated. As anticipated, the degree of bacterial inactivation increased
as a function of water availability up to a certain limit. This is probably because with increased
volume of immersion water, more pathogens are released onto the seed surface or into the
surrounding water with maximum release at an immersion volume of 3 ml. Charkowski et al.
(2001) reported that bacteria inside seeds may be internalized and are only released as the seeds
imbibe water, swell and expose previously sequestered regions of the seeds. As this happens, the
pathogen might become more prone to inactivation or disruption by high pressure. However,
unlike pathogen inactivation, the ability of seeds to germinate after pressure-treatment at 600
MPa was not significantly different when water was added to varying ratios. Since pressurization
of dry seeds impaired their ability to germinate, we can deduce that for high viability retention of
alfalfa seeds, presence of water during pressure treatment is much more critical than the actual
volume of water added. This compares well with the study undertaken by Wuytack et al. (2003)
who showed that garden cress seeds were found to achieve 100% germination rate when seeds
(1g) was pressure-treated with 0.25 ml of sterile DI water or a water to seed ratio of 1:4.
Moreover, Penas et al. (2008) showed that HHP process lowered the germination rates of alfalfa
and mung-bean seeds when wet/soaked seeds were pressure-treated in the absence of water
immersion. Hence in reconciling our findings with those of other authors, we can infer that

82
presence of water during pressure treatment was critical for seed viability. Pressurization of
soaked seeds without submerging them in water is likely to cause more structural damage to the
integrity of the seeds as a result of softening of their seed coat.
Whilst 600 MPa for 2 min was able to achieve a ~5 log reduction with seeds inoculated
with ~109 CFU/g, seeds inoculated with a lower E. coli level (~105 CFU/g) underwent only 3.7
log reductions. Indeed, the extent of microbial inactivation achieved at a particular pressure
treatment depends on a number of interacting factors, including the initial number of organisms
(Palou et al., 1999). It is obvious that with a high inoculation level, the pressure inactivation
effect was more appreciable, hence studies with a realistic contamination level was also needed.
Pressure inactivation of E. coli O157:H7 at 600 MPa for 2-6 min was correlated with treatment
time with a maximum inactivation at 6 min. However, survivors were detected by enrichment at
treatment times of > 6 minutes such that even 20 minutes at 600 MPa could not result in
complete inactivation. Generally, an increase in pressure increases microbial inactivation.
However, increasing the process time does not necessarily increase the lethal effect as noted by
Palou et al. (1999). When seeds were treated at 650 MPa, the shape of the survival curve of the
pathogen was very similar to the one at 600 MPa with a rapid initial drop in bacterial counts.
Indeed, there have been many reports showing that the inactivation is not always linear
(Earnshaw et al., 1995; Palou et al., 1999; Chen et al., 2007). It is actually not uncommon to find
that the curve showing log of survivors versus treatment time is concave with a rapid initial
decrease in log of survivors followed by a tailing effect, where there is essentially no further
inactivation as treatment time increases. Such inactivation curves have also been found with

83
other species such as Salmonella, Listeria and Yersinia enterocolitica (Metrick et al., 1989;
Earnshaw et al., 1995; Isaacs et al., 1995; Patterson et al., 1995; Chen and Hoover, 2003). These
tailing effects have been reported in thermal resistance studies, but the tails appear to be more
prominent with pressure treatment. It should be noted that the highest temperatures reached
during pressurization were approximately 35°C and 36°C for the 600 and 650 MPa treatments,
respectively. These relatively mild temperatures were not expected to contribute to the lethality
of the process.
E. coli O157:H7 is known to be a highly baro-resistant bacterium (Palou et al., 1999).
However, a combination of factors may also compound its resistance. It is likely that after the
seeds were inoculated and stored, the inoculated pathogen attached itself firmly to the seed
surfaces, underneath it or even permeated into the internal tissues via the hilum or the micropyle
of the seeds. E. coli O157:H7 is known to express multiple fimbrial and non-fimbrial adhesions
which may be involved in adhesion to surfaces (Torres et al., 2005). For example curli-
expressing thin aggregative fimbriae have been reported in E. coli O157:H7 to enable them to
bind to inert and abiotic surfaces. E. coli O157:H7 may form biofilms on the seed surfaces
which makes it more resistant to processing conditions. Severe pressure treatments were
therefore needed to eliminate this pathogen from seeds. In this study, a treatment of 650 MPa for
15 min at 20°C consistently achieved complete inactivation. This therefore demonstrates that
excessively high pressure magnitude in conjunction with long holding time is required to achieve
100% lethality. However, this extreme process condition did not have any deleterious effect on

84
the seed germination potential demonstrating therefore that HHP can be a highly feasible
technology to decontaminate seeds.
Conclusion
Decontamination of seeds prior to sprouting is critical in obtaining a safe product.
Although several decontamination methods have been proposed, these methods have proved to
be unsatisfactory. Surface irregularities, physical damage (e.g cracks, wrinkles, missing testa
parts) to seeds and the presence of pathogen bacterial biofilms on seed surfaces reduce the
effectiveness of chemical sanitizers. The presence of cracks and openings in the seed coat may
also provide a preferential pathway for entry of pathogens into the seed and may therefore
enhance the survival of pathogens inside the seed. Once pathogens are entrapped inside the seed,
targeting them becomes extremely difficult. In this study, HHP treatment of 650 MPa for 15 min
at 20°C was able to completely eliminate a > 5 log of E. coli O157:H7 on alfalfa seeds without
impairing the germinability of seeds. Since the initial pathogen contamination levels in seeds are
typically very low, < 1 MPN/g seed (Inami et al., 2001; Stewart et al., 2001), a treatment giving a
5-log reduction should be adequate.

85
References Ariefdjohan, M.W., Nelson, P.E., Singh, R.K., Bhunia, A.K., Balasubramaniam, V.M., Singh, N., 2004. Efficacy of high hydrostatic pressure treatment in reducing Escherichia coli O157:H7 and Listeria monocytogenes in alfalfa seeds. Journal of Food Science 69, 117-126. Barak, J.D., Delwiche, M.J., Sananikone, K., 2005. Comparison of primers for the detection of pathogenic Escherichia coli using real-time PCR. Letters in Applied Microbiology 41, 112-118. Beuchat, L.R., 1996. Pathogenic microorganisms associated with fresh produce. Journal of Food Protection 59, 204-216. Bhagwat, A.A., Chan, L., Han, R., Tan, J., Kothary, M., Jean-Gilles, J., Tall, B.D., 2005. Characterization of enterohemorrhagic Escherichia coli strains based on acid resistance phenotypes. Infection and Immunity 73, 4993-5003. Charkowski, A.O., Sarreal, C.Z., Mandrell, R.E., 2001. Wrinkled alfalfa seeds harbor more aerobic bacteria and are more difficult to sanitize than smooth seeds. Journal of Food Protection 64, 1292-1298. Chen, H., Hoover, D.G., 2003. Pressure inactivation kinetics of Yersinia enterocolitica ATCC 35669. International Journal of Food Microbiology 87, 161-171. Chen, H., Hoover, D.G., Kingsley, D.H., 2005. Temperature and treatment time influence high hydrostatic pressure inactivation of feline calicivirus, a norovirus surrogate. Journal of Food Protection 68, 2389-2394. Chen, H., 2007. Use of linear, Weibull, and log-logistic functions to model pressure inactivation of seven food-borne pathogens in milk. Food Microbiology 24, 197-204. Earnshaw, R.G., Appleyard, J., Hurst, R.M., 1995. Understanding physical inactivation processes: Combined preservation opportunities using heat, ultrasound and pressure. International Journal of Food Microbiology 28, 197-219. Farkas, D.F., Hoover, D.G., 2000. High-pressure processing. Journal of Food Science Supplement 65, 47-64. Fett, W.F., 2006. Interventions to ensure the microbial safety of sprouts. In: Sapers, G.M., Gorny, J.R., Yousef, A.E. (Eds.), Microbiology of Fruits and Vegetables. Taylor and Francis Group, LLC, Boca Raton, FL., pp. 187-210.

86
Hitchins, A.D., Feng, P., Watkins, W.D., Rippey, S.R., Chandler, L.A. 1995. Escherichia coli and the Coliform Bacteria. U.S. FDA Bacteriological Analytical Manual, Chapter 4. AOAC International, Gaithersburg, Maryland, U.S.A. Hu, H., Churey, J.J., Worobo, R.W., 2004. Heat treatments to enhance the safety of mung bean seeds. Journal of Food Protection 67, 1257-1260. Ibrahim, S.A., Salameh, M.M., Phetsomphou, S., Yang, H., Seo, C.W., 2004. Application of caffeine, 1,3,7-trimethylxanthine, to control Escherichia coli O157:H7. Food Chemistry 99, 645-650. Inami, G.B., Lee, S.M.C., Hogue, R.W., Brenden, R.A., 2001. Two processing methods for the isolation of Salmonella from naturally contaminated alfalfa seeds. Journal of Food Protection 64, 1240-1243. Isaacs, N.S., Chilton, P., Mackey, B., 1995. Studies on the inactivation by pressure of micro-organisms. In: Ledward, D.A., Johnston, D.E., Earnshaw, R.G., Hasting, A.P.M. (Eds.), High Pressure Processing of Foods, Nottingham University Press, Nottingham, U.K., pp.65-79. Kingsley, D.H., Chen, H., 2008. Aqueous matrix compositions and pH influence virus inactivation by high pressure processing. Journal of Food Protection. In press. Metrick, C., Hoover, D.G., Farkas, D.F., 1989. Effects of high-hydrostatic pressure on heat-resistant and heat-sensitive strains of Salmonella. Journal of Food Science 54, 1547. Michel, M., Autio, K., 2001. Effects of high pressure on protein- and polysaccharide-based structures. In: Hendricks, M.E.G., Knorr, D. (Eds.), Ultra High Pressure Treatments of Foods. Kluwer Academic/Plenum Publishers, New York, NY, pp. 189-210. Mundt, J.O., Hinkle, N.F., 1976. Bacteria within ovules and seeds. Applied and Environmental Microbiology 32, 694-698. NACMCF (National Advisory Committee on Microbiological Criteria for Foods), 1999. Microbiological safety evaluations and recommendations on sprouted seeds. International Journal of Food Microbiology 52, 123-153. Ogawa, T., Nakatani, A., Matsuzaki, H., Isobe, S., Isshike, K., 2000. Combined effects of high hydrostatic pressure, temperature, and the addition of allyl-isothiocyanate on inactivation of Escherichia coli. Journal of Food Protection 63, 884-888. Oxen, P., Knorr, D., 1993. Baroprotective effects of high solute concentrations against inactivation of Rhodotorula rubra. LWT- Food Science and Technology, 26, 220-223.

87
Palou, E., Lopez-Malo, A., Barbosa-Canovas, G.V., Welti-Chanes, J., Swanson, B.G., 1997. Effect of water activity on high hydrostatic pressure inhibition of Zygosaccharomyces bailii. Letters in Applied Microbiology 24, 417-420. Palou, E., Lopez-Malo, A., Barbosa-Canovas, G.V., Swanson, B.G., 1999. High pressure treatment in food preservation. In: Rahman, M.S. (Ed.), Handbook of food preservation, Marcel Dekker, New York, NY, pp. 533-576. Patterson, M.F., Quinn, M., Simpson, R., Gilmour, A., 1995. Effects of high pressure on vegetative pathogens. In: Ledward, D.A., Johnston, D.E., Earnshaw, R.G., Hasting, A.P.M. (Eds.), High Pressure Processing of Foods, Nottingham University Press, Nottingham, U.K., pp.47-63. Penas, E., Vidal-Valverde, C., Frias, J., Gomez, R., 2008. Application of high-pressure treatment on alfalfa (Medicago sativa) and mung bean (Vigna radiata) seeds to enhance the microbiological safety of their sprouts. Food Control 19, 698-705. Rajkowski, K.T., Thayer, D.W., 2001. Alfalfa seed germination and yield ratio and alfalfa sprout microbial keeping quality following irradiation of seeds and sprouts. Journal of Food Protection 64, 1988-1995. Sharma, R.R., Fett, W.F., Beuchat, L.R., Demirci, A., 2003. Application of ozone for inactivation of Escherichia coli O157:H7 on inoculated alfalfa sprouts. Journal of Food Processing and Preservation 27, 51-64. Smelt, J.P., Hellemons, J.C., Patterson, M., 2001. Effects of high pressure on vegetative microorganism. In: Hendricks, M.E.G., Knorr, D. (Eds.), Ultra High Pressure Treatment of Foods, Kluwer Academic/Plenum, New York, NY., pp. 55-76. Stewart, D.S., Reineke, K.F., Ulaszek, J.M., Tortorello, M.L., 2001. Growth of Salmonella during sprouting of alfalfa seeds associated with salmonellosis outbreaks. Journal of Food Protection 64, 618-622. Taormina, P.J., Beuchat, L.R., 1999. Comparison of chemical treatments to eliminate enterohemorrhagic Escherichia coli O157:H7 on alfalfa seeds. Journal of Food Protection 62, 318-324. Taormina, P.J., Beuchat, L.R., 1999. Behavior of enterohemorrhagic Escherichia coli O157:H7 on alfalfa sprouts during the sprouting process as influenced by treatments with various chemicals. Journal of Food Protection 62, 850-856.

88
Torres, A.G., Matthysse, A.G., Langley, W., Jeter, C., 2005. Differential binding of Escherichia coli O157:H7 to alfalfa, human epithelial cells, and plastic is mediated by a variety of surface structures. Applied and Environmental Microbiology 71, 8008-8015. U.S. Food and Drug Administration, 1999. Guidance for industry: sampling and microbial testing of spent irrigation water during sprout production. Docket no. 99D-4488. Available at: http://vw.cfsan.fda.gov/~dms/sprougd1.html. Utlee, A., Gorris, L.G.M., Smid, E.J., 1998. Bacterial activity of carvacrol towards the food-borne pathogen Bacillus cereus. Journal of Applied Microbiology 85, 211-218. Waddell, J.M, Troxell, T.C., 2000. Safer processing of sprouts, U.S. FDA and the California Department of Health Services, Food and Drug Branch, available at http://vm.cfsan.fda.gov/˜dms/sprouvid.html Weissinger, W.R., Beuchat, L.R., McWatters, K.H., 2001. Evaluation of volatile chemical treatments for lethality to Salmonella on alfalfa seeds and sprouts. Journal of Food Protection 64, 442-450. Wuytack, E.Y., Michiels, C.W., Meersseman, K., Diels, A.M., 2003. Decontamination of seeds for sprout production by high hydrostatic pressure. Journal of Food Protection 66, 918-923.
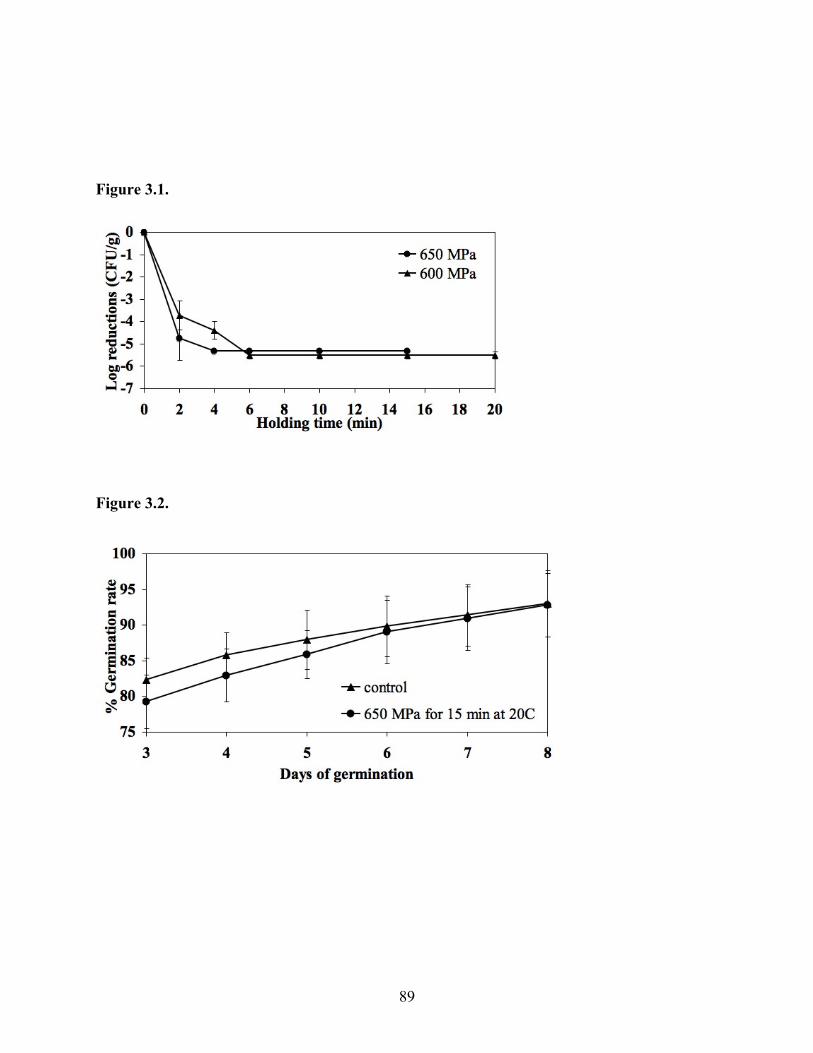
89
Figure 3.1.
Figure 3.2.

90
Table 3.1 Effect of pressure treatment and seed wetness on the percentage germination rate of mung bean seeds. Data are the means of three replicates ± one standard deviation. Days of Pressure-treatment at various pressure levels germination Control State 200 MPa 300 MPa 400 MPa 500 MPa 600 MPa 3 94.4±11.8 a Dry 5.4±6.2 b 1.4±0.6 b 6.9±9.2 b 2.0±1.0 b 4.9±1.6 b Wet 77.7±3.5 ac 72.7±7.0 c 76.2±4.0 c 76.0±4.6 c 72.4±2.6 c 4 99.2±2.0 a Dry 5.4±6.2 b 1.4±0.6 b 8.6±12.2 b 2.0±1.0 b 4.9±1.6 b Wet 89.5±5.3 ac 80.6±3.5 c 84.2±3.0 c 86.2±0.4 ac 85.6±2.7 ac 5 99.6±1.0 a Dry 5.4±6.2 b 1.7±1.2 b 9.7±14.0 b 2.7±1.6 b 4.9±1.6 b Wet 91.9±4.0 ac 83.9±5.0 c 90.0±2.8 ac 88.5±1.8 ac 86.6±1.7 ac 6 99.6±1.0 a Dry 5.4±6.2 b 1.7±1.2 b 9.7±14.0 b 2.7±1.6 b 4.9±1.6 b Wet 93.8±3.1 a 84.9±5.0 a 92.7±2.8 a 91.1±1.2 a 87.2±1.8 a 7 99.8±0.5 a Dry 5.4±6.2 b 1.7±1.2 b 9.7±14.0 b 2.7±1.6 b 4.9±1.6 b Wet 94.5±2.3 a 85.2±5.1 a 93.3±2.5 a 91.8±1.7 a 87.2±1.8 a 8 100.0±0.0 a Dry 5.4±6.2 b 1.7±1.2 b 9.7±14.0 b 2.7±1.6 b 4.9±1.6 b Wet 94.5±2.3 a 86.2±4.4 a 93.3±2.5 a 92.4±1.7 a 88.1±1.7 a
Values for the same day of germination (dry or wet state) followed by the same letter are not significantly different (P > 0.05). Table 3.2 Effect of pressure treatment and seed wetness on the percentage germination rate of alfalfa seeds. Data are the means of three replicates ± one standard deviation. Days of Pressure-treatment at various pressure levels germination Control State 200 MPa 300 MPa 400 MPa 500 MPa 600 MPa 3 94.4±11.8 a Dry 61.0±17.6 a 58.9±18.9 a 53.2±15.1 a 63.0±23.6 a 47.1±12.8 a Wet 71.9±24.9 a 71.4±17.9 a 68.4±24.7 a 71.8±22.7 a 92.5±1.2 a 4 99.2±2.0 a Dry 71.4±13.3 a 70.1±13.9 a 64.1±13.0 a 74.0±21.3 a 65.9±14.2 a Wet 84.1±13.0 a 80.0±11.6 a 78.4±18.5 a 83.9±17.7 a 94.8±0.6 a 5 99.6±1.0 a Dry 75.6±12.8 a 75.4±12.9 a 71.8±13.5 a 77.6±19.3 a 75.7±13.5 a Wet 87.8±9.3 a 85.9±7.0 a 84.4±13.9 a 90.8±11.6 a 97.1±0.9 a 6 99.6±1.0 a Dry 80.7±13.2 a 82.8±7.8 a 77.5±11.2 a 84.6±14.2 a 83.8±11.1 a Wet 93.3±6.0 a 89.7±7.0 a 88.4±10.7 a 94.0±7.1 a 98.2±1.3 a 7 99.8±0.5 a Dry 84.2±15.7 a 85.8±7.0 a 82.3±10.2 a 86.8±13.2 a 87.3±10.0 a Wet 96.2±3.4 a 93.4±8.0 a 92.0±8.7 a 95.5±5.3 a 98.2±1.3 a 8 100.0±0.0 a Dry 85.4±15.4 a 90.5±7.8 a 87.5±6.9 a 89.5±11.5 a 90.5±7.9 a Wet 97.2±2.5 a 94.7±7.4 a 94.1±7.5 a 96.5±4.7 a 98.8±1.2 a
Values for the same day of germination (dry or wet state) followed by the same letter are not significantly different (P > 0.05).

91
Table 3.3 Effect of volume of water immersion during pressure treatment at 600 MPa for 2 min at 20°C on the baro-inactivation of E. coli O157:H7 and the percentage germination rate of seeds. Data are the means of three replicates ± one standard deviation.
Water Log Percentage germination rate at different days Volume
(ml) reductions 3 4 5 6 7 8 0 -0.4 ± 0.2 a
47.1±12.8 a 65.9 ±14.2 a 75.7 ± 13.5 a 83.8 ± 11.1 a 87.3 ± 10.0 a 90.5 ± 7.9 a
1 -2.3 ± 0.1 b 86.4± 3.3 b 91.8± 3.2 b 95.5 ± 2.6 a 97.8 ± 3.0 a 98.4 ± 1.9 a 99.1 ± 1.6 a
2 -4.3 ± 0.2 c 85.0± 4.5 b 90.7± 4.1 b 94.2 ± 2.3 a 97.2 ± 1.7 a 98.6 ± 1.3 a 99.2 ± 0.8 a
3 -4.4 ± 0.4 c 87.0± 2.5 b 91.6± 3.2 b 95.2 ± 1.6 a 96.6 ± 1.4 a 98.0 ± 1.8 a 98.5 ± 1.4 a
5 -4.4 ± 0.3 c 86.3± 3.8 b 91.6± 2.9 b 94.8 ± 1.1 a 96.5 ± 0.4 a 96.8 ± 1.0 a 96.8 ± 1.0 a Values in the same column followed by the same letter are not significantly different (P > 0.05).

92
Chapter 4
Factors affecting the efficacy of pressure inactivation of Escherichia coli O157:H7
on alfalfa seeds and seed viability
Published in Int. J. Food Microbiol. 2009. 131, 218-223

93
Abstract
The application of high hydrostatic pressure technology as a seed decontamination
technology was evaluated. Alfalfa seeds inoculated with approximately 105 CFU/g of
Escherichia coli O157:H7 were subjected to oscillatory pressure treatments at 600 MPa and
20°C for up to five cycles with a holding time of 2 min/cycle. However, oscillatory
pressurization was not able to eliminate E. coli O157:H7. The application of pressure
treatment at 600 MPa for 2 min at 20°C in the presence of chemicals such as calcium
hypochlorite, calcium hydroxide, lactic acid or sodium acid sulfate was subsequently
investigated and it was demonstrated that this “multiple hurdle” approach was unable to
decontaminate alfalfa seeds. Soaking seeds prior to pressure treatment was found to play a
critical role on enhancing the pressure inactivation of E. coli O157:H7; seeds soaked in water
for 60 min followed by treatment at 600 MPa for 2 min at 20°C were decontaminated and had
a germination rate of 91% which was 4% lower than that of the untreated seeds (not
statistically significant, P > 0.05). It was further demonstrated that a process involving
soaking seeds in water for ≥ 10 min followed by treatment at 600 MPa for 15 min at 20°C
was equally effective with respect to E. coli O157:H7 elimination and viability retention of
the seeds.

94
Introduction
Numerous outbreaks of food-borne diseases linked to consumption of fresh fruits and
vegetables clearly point to the fact that these commodities constitute a serious hazard to
public health. Seed sprouts are regarded by consumers as a health food because they are low
in fat and calories, high in fiber and are rich in anti-cholesterolemic and anti-carcinogenic
constituents (Kurtzweil, 1999). However, sprouted seeds continue to be implicated in
outbreaks of food-borne illness and hence are considered a significant food safety risk
(Anonymous, 1999; Kumar et al., 2006). Since 1995, there have been three outbreaks
associated with the consumption of alfalfa and clover sprouts contaminated with E. coli O157
in the United States (Fett, 2002; Ferguson et al., 2005). In the majority of the illness
outbreaks, the seeds used for sprout production were proven to be the most likely source of
human pathogens (Anonymous, 1999).
Alfalfa sprouts are typically eaten raw and therefore intervention treatments must be
either applied to the seeds or to the germinated sprouts. Since sprout-directed treatments could
be more problematic because of the delicate nature of the sprouts, more research attention has
been focused on alfalfa seed decontamination as opposed to treatment on alfalfa sprouts (Lang
et al., 2000). In response to the urgent need to curtail sprout-related outbreaks and to protect
public safety, the U.S. Food and Drug Administration and the State of California are strongly
recommending the treatment of alfalfa seeds with a solution of 20,000 ppm free chlorine from
calcium hypochlorite. This treatment is currently considered the “benchmark” and is also
recommended by the National Advisory Committee on Microbiological Criteria for Foods
(Anonymous, 1999). However, it has been shown that this method may not be robust enough

95
for seed decontamination. It reduces, but does not eliminate pathogens on seeds and
consequently outbreaks continue to occur (Brooks et al., 2001; Proctor et al., 2001). As a
result, extensive research is currently ongoing to investigate the ability of various physical,
chemical and biological preventive strategies to decontaminate seeds from foodborne
pathogens.
High hydrostatic pressure (HHP) processing is a non-thermal food preservation
technology. One of the most important effects of HHP processing is its ability to destroy or
reduces the number of foodborne pathogens with minimum impact on food sensory and
functional qualities. Previous researchers investigating the application of HHP (Ariefdjohan
et al., 2004; Wuytack et al., 2003) to decontaminate alfalfa and mung bean seeds have
reported varying degrees of success due to the inability of high pressure to eliminate
pathogenic microorganisms from seeds with minimal impact on the seeds germinability.
Since these seeds are destined for sprouting, retention of their seed viability is critical to
ensure that pressure-treated seeds can consistently meet the yield anticipated by sprouters. A
previous study in our laboratory demonstrated that continuous high pressure treatment at 600
MPa for 20 min at 20°C achieved a ~5 log reduction but could not eliminate E. coli O157:H7
(105 CFU/g) on alfalfa seeds (Neetoo et al., 2008). Therefore the overall aim of this study was
to develop a HHP process to decontaminate alfalfa seeds inoculated with a ~ 5 log CFU/g of
E. coli O157:H7 with minimal adverse impact on the seeds germination potential. The
specific objectives were to determine the effect of (1) oscillatory HHP treatments, (2) the
combined effect of HHP with antimicrobial agents and (3) the application of HHP on pre-
soaked seeds for varying holding times on seed decontamination and viability.

96
Materials and Methods
Bacterial Strains
E. coli O157:H7 strains 1730, 250, 251, Cider and J58, adapted to grow in tryptic soy
broth plus 0.6% yeast extract (Difco Laboratories, Sparks, MD, USA) supplemented with 50
µg/ml of nalidixic acid (Fisher Scientific, Hampton, NH, USA) (TSBYE-N) were used. The
five-strain cocktail of E. coli O157:H7 was prepared as described by Neetoo et al. (2008).
Inoculation of seeds
The cocktail (10 ml) was mixed with 100 ml of sterile 0.1% peptone water (Fisher).
Unscarified and undamaged alfalfa seeds (100 g), obtained from International Specialty
Supply (Cookeville, Tenn., USA), were added to the cell suspension (21± 1°C) and gently
stirred for 5 min. The seeds were separated from the cell suspension by pouring the mixture
over a double layer of cheesecloth supported by a wire screen and dried inside a bio-safety
hood at room temperature (21 ± 1°C) for 24 h to a final aw of 0.622. Dried seeds with an
approximate inoculation level of 105 CFU/g of E. coli O157:H7 were placed in sterile pouches
and stored at 4°C until needed.
Pressure treatment
Two g of inoculated seeds and 3 ml of sterile DI water were placed in a 3-mil thick
pouch (Nylon/Polyethylene, Koch Supplies, Kansas City, MO). To avoid leakage during
pressure-treatment, each sample pouch was placed in a larger pouch of an 8-mil thick PVC
plastic (McMaster-Carr, Elmhurst, IL) and heat-sealed. HHP treatment of samples was carried
out using a high-pressure unit with temperature control (Model Avure PT-1, Avure

97
Technologies, Kent, WA). All the experiments were conducted at 20°C (initial seed sample
temperature prior to pressure treatment) using water as a hydrostatic medium. The
temperature of the water-bath surrounding the pressure chamber during pressurization was
monitored through K-type thermocouples. The temperature and pressure data were recorded
every 2 s (DASYTEC USA, Bedford, NH). The pressure-come-up rate was approximately 22
MPa/s. The pressure-release was < 4 s. Pressurization time reported in this study did not
include the pressure come-up or release times. Oscillatory pressurization was performed by
alternating atmospheric pressure (0.1 MPa) and high pressure (600 MPa) with 1, 2, 3, 4 or 5
cycles and a holding time of 2 min/cycle.
Microbiological analysis
Pouches containing treated seeds were cut open aseptically. The seed mixture
consisting of the seeds and immersion water was poured into a stomacher bag to which 8 ml
of sterile 0.1% peptone water was added and subsequently stomached for 2 min at 260 rpm
(Seward 400 Stomacher, Seward Medical Co., London, U.K.). The seed slurry was serially
diluted in sterile 0.1% peptone and surface-plated in duplicate on tryptic soy agar with 0.6%
yeast extract (Difco Laboratories, Sparks, MD, USA) supplemented with nalidixic acid to a
final concentration of 50 µg/ml (TSAYE-N). TSAYE-N plates were incubated at 35ºC for 3
days. Presumptive colonies of E. coli O157:H7 formed on the plates were enumerated. One or
two colonies were picked at an initial stage in the study and confirmed for E. coli O157:H7
identification using either a BAX® system for screening E. coli O157:H7 PCR assay
(Qualicon-DuPont, Wilmington, DE, USA) or Rapid E. coli O157:H7 Test Methods
(Strategic Diagnostics Inc., Newark, DE, USA).

98
Combined effect of HHP and antimicrobial treatments on inactivation of E. coli
O157:H7 on alfalfa seeds
The combined application of HHP and antimicrobial treatments on seed
decontamination was investigated. Inoculated seeds were either (a) soaked in antimicrobial
solutions for varying durations and then subjected to a 2 min-pressure treatment or (b) directly
pressure-treated in antimicrobial solutions (without prior soaking) for extended exposure
times. In the first approach, 2 g of inoculated seeds were soaked in 20 ml of DI-water
(control), calcium hypochlorite (Acros Organics, N.J.) solutions providing approximately 200,
1000, 2,000, or 20,000 ppm of free chlorine as determined using chlorine test strips (Fisher),
1% calcium hydroxide (Reheis Inc., Midlothian, TX) with or without 1% Tween 80 (Fisher),
lactic acid (1, 2 or 5%) (Fisher), or sodium acid sulfate (0.05, 0.10 or 0.20%) (Jones-
Hamilton, Walbridge, OH) for 5, 10 or 15 min at room temperature (21 ± 1°C). The
concentration unit was defined as %w/v. The calcium hypochlorite solutions were prepared in
a sterile 0.05 M potassium phosphate buffer solution with pH of 6.8. After soaking, the excess
water or chemical solutions was decanted and the seeds were placed into a sterile plastic
pouch followed by the addition of 3 ml of the respective treatment solutions and treated at 600
MPa for 2 min at 20°C. In the second approach, 2 g of the inoculated seeds were placed into
sterile pouches and pressure-treated at 600 MPa and 20°C whilst submerged in 3 ml of
various antimicrobial solutions for varying holding times (5, 10 or 15 min). The antimicrobial
solutions included 2,000 and 20,000 ppm calcium hypochlorite, 1% calcium hydroxide with
or without 1% Tween 80 and 1, 2 and 5% lactic acid. Pouches containing treated seeds were
cut open aseptically. Seed samples were rinsed with 100 ml of sterile DI-water to remove any

99
residues of the chemical solutions and then poured into a stomacher bag to which 8 ml of
sterile 0.1% peptone water was added and subsequently stomached for 2 min. The seed slurry
was directly enriched in 90 ml of TSBYE-N and incubated for 48 h at 35ºC to allow
resuscitation of sub-lethally injured cells. Samples were streaked onto Sorbitol MacConkey
agar (Difco Laboratories, Sparks, MD, U.S.A.) plates supplemented with 50 µg/ml of
nalidixic acid. After 24 h incubation, presence of growth exhibiting morphological and
biochemical characteristics typical of E. coli O157:H7 were determined by visually inspecting
the plates. Colonies were confirmed to be E. coli O157:H7 using Rapid E. coli O157:H7 Test
Methods (Strategic Diagnostics Inc., Newark, DE, USA).
Effect of soaking time in water prior to pressure treatment on inactivation of E. coli
O157:H7 on alfalfa seeds and seed viability
Two g of inoculated seeds was soaked in sterile DI-water for 0 (without soaking), 30,
60, 90, 120, 180, 240 or 300 min prior to being treated at 600 MPa for 2 min at 20°C. Seeds
were soaked either in a limiting volume (3 ml) or in a large excess (20 ml) of water to
determine whether the soaking time as well as the volume of water available were critical
factors in the pressure inactivation of E. coli O157:H7 on seeds. For seeds soaked in 3 ml of
water, the 2 g inoculated seeds and 3 ml water were directly placed into a pouch and soaked
for varying durations before being subjected to the pressure treatment. For seeds soaked in 20
ml of water, the excess water was subsequently decanted and seeds were placed into a pouch
in the presence of 3 ml of fresh sterile DI water and pressure-treated. Samples were then
microbiologically assayed post-treatment as described previously. Soaked controls were
additionally set up by immersing inoculated seeds in 20 ml of sterile DI water for 30, 60, 90,

100
120, 180, 240 or 300 min before determining the remaining populations of E. coli O157:H7
on the seeds.
To determine the effect of soaking duration prior to pressure treatment on the seeds
germination potential, 2 g of un-inoculated seeds were soaked in 20 ml of water for 0, 30, 60,
90, 120 or 180 min. The water was then decanted and the seeds introduced into a pouch to
which 3 ml of DI-water was added. The samples were treated at 600 MPa for 2 min at 20°C.
After HHP, pressure-treated seeds and untreated seeds (control) were soaked in DI-water for a
total soaking time of 3 hrs (including the soaking times before and after the pressure
treatment). One hundred seeds were drawn from the soaked seeds and spread evenly on pieces
of wet paper towels on a plastic rack, which in turn was placed into a water-filled bucket to
provide a moist environment for the seeds. The water level was maintained below the seeds’
level. The bucket was kept at room temperature (21 ± 1°C) for 8 days (as suggested by the
seeds provider) and misted daily. The bucket was covered loosely with a piece of plastic film
to allow exchange of air between the inside and outside of the bucket. The seeds were visually
evaluated for sprouting 3 to 8 days after setting up the germination system.
To determine the rate of water absorption by seeds, 2 g of un-inoculated seeds were
immersed in 20 ml of water at room temperature. Soaked seeds were weighed and their
weight gain recorded at regular time intervals for up to 19 hrs.
Effect of reduced soaking time and extended pressure holding time on the inactivation of
E. coli O157:H7 and seed viability
Two g of inoculated seeds were soaked in 20 ml of water for 10 or 15 min. The water
was later decanted and seeds mixed with 3 ml of sterile DI-water, packaged and treated at 600

101
MPa and 20°C for 5, 10 or 15 min. Samples were then microbiologically assayed as described
previously and enriched for the detection of survivors. For the determination of seed
germination, 2 g of un-inoculated seeds were soaked for 10 or 15 min and treated at 600 MPa
for 15 min at 20°C. One hundred seeds were drawn from the samples and assayed for
germination as described previously.
Statistical analysis
All experiments were replicated at least three times. Where appropriate, statistical
analyses were conducted using Minitab® Release 15 (Minitab Inc., University Park, PA,
USA). One-way analysis of variance (ANOVA) and Tukey’s one-way multiple comparisons
were used to determine differences in the populations of E. coli O157:H7 recovered on treated
alfalfa seeds as well as differences in the germination percentage of seeds. Significant
differences were considered at the 95% confidence level (P < 0.05).
Results and Discussion
Effect of oscillatory pressurization on the inactivation of E. coli O157:H7 on alfalfa seeds
Oscillatory pressure treatment was investigated to determine whether it could be used
to enhance pressure inactivation since continuous high pressure treatment at 600 MPa for 20
min at 20°C could not eliminate E. coli O157:H7 (105 CFU/g) on alfalfa seeds (Neetoo et al.,
2008). Results for the oscillatory HHP treatments are summarized in Figure 4.1. The degree
of pressure inactivation was found to vary as a function of the number of cycles applied,
achieving a maximum reduction of 3.7 logs after 5 cycles for a total holding time of 10 min at
600 MPa. Contrary to findings garnered by other researchers, oscillatory pressure treatments
did not confer a significant advantage in the inactivation of E. coli O157:H7. Palou et al.

102
(1998) evaluated the inactivation of Zygosaccharomyces bailii by oscillatory pressure and
continuous pressure treatments and found that oscillatory pressure treatments increased
pressure inactivation. Aleman et al. (1996) also reported greater reductions of Saccharomyces
cerevisiae when oscillatory pressure treatments were applied. Enhancement in oscillatory
pressure inactivation has also been reported for spores (Hayakawa et al., 1998). It is thought
that the effectiveness of oscillatory HHP treatments can be attributed to spore burst which can
be promoted by increased spore wall permeability at high pressures (Palou et al. 1998).
However, our results are comparable to those reported by Kingsley et al. (2006) for the
pressure inactivation of hepatitis A virus (HAV). Oscillatory high pressure processing for 2,
4, 6, and 8 cycles at 400 MPa did not considerably enhance the inactivation of HAV as
compared with continuous high pressure application.
Combined effect of HHP and antimicrobial treatments on inactivation of E. coli
O157:H7 on alfalfa seeds
Since oscillatory pressurization with as many as 5 cycles could not eliminate 105
CFU/g of E. coli O157:H7, a “multiple hurdle approach” was investigated whereby the
simultaneous application of antimicrobials and HHP were evaluated on inoculated alfalfa
seeds. Previous research has shown that washing or soaking in antimicrobial agents can
reduce the microbial load in seeds (Jaquette et al., 1996; Piernas et al., 1997). The drawback
associated with these chemical treatments however is that high antimicrobial concentrations
are usually required to demonstrate any appreciable effect and the treatments can not
eliminate pathogens. In this experiment, the concerted application of HHP and chemical
treatments was investigated. Calcium hydroxide was included in this study as an alternative to

103
chlorinated sanitizers while lactic acid and sodium acid sulfate were studied because of their
acidulating property, which was thought to enhance pressure inactivation of bacterial
pathogens. Since we were interested in determining whether the application of HHP in
conjunction with chemical treatments would be able to achieve 100% lethality or not, samples
were analyzed qualitatively for presence/absence of survivors after enrichment in non-
selective media. Soaking seeds in the different chemical solutions at low, medium or high
concentrations prior to pressure treatment did not achieve 100% kill of the pathogen as
survivors were consistently detected across all treatments in all three trials. Since the 2-min
pressure holding time did not bring about elimination of E. coli O157:H7, seeds were then
pressure-treated with selected antimicrobials consisting of calcium hypochlorite, calcium
hydroxide or lactic acid with an extended pressure holding time of up to 15 min. Regardless
of the nature or the level of antimicrobials used, survivors were still detected post-enrichment.
It is believed that a small population of E. coli O157:H7 was particularly resistant to
pressure treatments. Indeed, other authors have shown that vegetative bacteria such as E. coli
O157:H7 can be very refractory to high pressure with pronounced “tails” on survival curves
(Chen, 2007). These pressure-resistant cells might be sub-lethally injured during a pressure
treatment and with subsequent appropriate conditions conducive to growth, these cells can
recover (Earnshaw et al., 1995). Hence sub-lethal damage and subsequent recovery can
present a significant problem for alfalfa sprouts that are eaten raw given that these recalcitrant
pathogenic cells can multiply during sprouting.
Past research by other authors has demonstrated that chemical sanitizers as stand-alone
treatments are not effective at eliminating E. coli O157:H7 from alfalfa seeds (Beuchat and

104
Scouten, 2002; Taormina and Beuchat, 1999; Weissinger and Beuchat, 2000). It was thus
hypothesized in this study that the simultaneous application of antibacterial agents and HHP
could provide a good opportunity for the chemicals to capitalize on the widespread cellular
and biochemical damages induced by pressure treatment. The fact that pressurization of the
seeds in the presence of very high concentration of these compounds could not eliminate E.
coli O157:H7 lends strong credence to the explanation that these pathogens are lodged in deep
cracks or crevices on the seed coat or even internalized. As a result, these chemicals cannot
permeate the seed coat to target those internalized cells within the time-frame investigated in
our study.
Microscopy of mung bean seeds has shown that although the seed surface is relatively
smooth, the stem scar is porous, allowing bacteria to penetrate into the seed. Microscopic
examination of alfalfa seeds has revealed similarities in the seed coat, both being smooth but
with areas capable of harboring pathogens, thus protecting them from aqueous sanitizers
(Delaquis et al., 1999). In addition to its topographic complexity, the surface of alfalfa seeds
is known to be covered with a waxy cuticle (cutin) and it is unlikely that aqueous solutions
can effectively wet these surfaces. Given the hydrophobic nature of alfalfa seeds, we
hypothesized that addition of a surfactant such as Tween 80 as an adjunct to 1% calcium
hydroxide could enhance permeation of the surfactant or the sanitizer into the seeds.
However, contrary to findings reported by Weissinger and Beuchat (2000), we found that the
application of surfactant during pressure treatment did not enhance the antimicrobial activity
of calcium hydroxide and was not effective in achieving complete lethality.

105
Effect of soaking time and immersion volume in water prior to pressure treatment on
inactivation of E. coli O157:H7 on alfalfa seeds
Since the combined application of antimicrobial treatments with HHP was
unsuccessful in achieving elimination of E. coli O157:H7, modification of the HHP strategy
to decontaminate seeds was then sought in an attempt to enhance the microbial safety of
alfalfa seeds. Seeds soaked in sterile DI water for up to 300 min (soaked controls) did not
undergo any significant reduction in the counts of E. coli O157:H7 with final populations
ranging from 4.7-5.3 log CFU/g. Table 4.3 shows that treatment at 600 MPa without prior
soaking brought about a 2.7-log reduction. Pre-soaking seeds in water was accompanied by a
greater extent of inactivation; in fact a direct relationship between the degree of inactivation
and soaking time were observed. When seeds were soaked in a limited volume of 3 ml of
water, pressure inactivation of the vegetative cells was strongly dependent on the soaking
time. This was likely due to the fact that longer soaking times allowed water to permeate
deeper into the cracks and crevices of seeds raising their local water activity and hence
enhancing pressure inactivation of cells trapped in these spaces. Seeds soaked for 120-180
min had no detectable counts although survivors were still detected post-enrichment.
However, soaking for ≥ 240 min followed by pressure treatment achieved elimination of the 5
log CFU/g initial load. Seeds were also soaked in a large excess of water (20 ml) such that the
volume of water for soaking would not be a limiting factor. The same trend was observed i.e
the longer the soaking time, the greater the degree of pressure inactivation. After seeds were
soaked for ≥ 60 min followed by pressure treatment, elimination of E. coli O157:H7 was
consistently achieved. Hence it can be concluded that both the soaking time and water volume

106
are critical factors in ensuring the efficacy of HHP to decontaminate alfalfa seeds.
In the light of these results, it can be inferred that in the presence of excess soaking
water, there was uninhibited penetration of water into the seeds such that an hour of
imbibition was adequate to allow water to access the deepest cracks or crevices of the seed
coat. In the presence of limiting volume of water, water uptake was slower; delaying the
consequent movement of water into the seed coat spaces. The results from this soaking study
reinforce the findings reported in Section 3.2 suggesting that the poor efficacy of HHP with
antimicrobial agents might be attributed to the sub-surface location of the pathogen on the
seed. It is thought the pre-soaking step is critical in raising the water activity of the seeds
which is known to have a bearing on the pressure inactivation of vegetative bacterial cells. It
is well documented that low water activity levels protect microorganisms against pressure
treatment (Oxen and Knorr, 1993; Palou et al., 1997; Kingsley and Chen, 2008).
Effect of soaking time in water prior to pressure treatment on the seed viability
To determine the effect of pre-soaking time on seed viability, the germination rate of
seeds soaked for varying durations before HHP was determined (Table 4.4). Soaking for ≤ 60
min followed by HHP still allowed seeds to retain their viability to a large extent. The process
of soaking seeds in 20 ml of water for 60 min followed by pressure treatment for 2 min
achieved a final germination yield of 91%, which was 4% lower than the control. This
difference in seed germination was not statistically significant (P > 0.05). However with
prolonged soaking of > 60 min, the effect of pressure treatment on the seeds’ germinability
became more severe. It is not clear what mechanisms are responsible for the progressively
lower germination yields but it is possible that denaturation of molecules such as enzymes

107
might have occurred during HHP following their activation during prolonged water
imbibition. It is also highly likely that during soaking, seeds imbibe water and as a result of
the increased moisture content, the seeds advance into an active physiological state whereby
metabolic changes associated with the initial stages of germination start to take place. The
effect of high pressure on the delicate early germinative stage of the seeds most probably
evoked negative changes in the microstructure of the seeds. Blaszczak et al. (2007) compared
the micro-structural and biochemical changes undergone in raw and germinated chickpea
seeds after pressure treatment. They demonstrated the occurrence of more pronounced
damage to the structural integrity (physical damage) and protoplasmic contents of cells
(biochemical damage) after germinated chickpea seeds were pressure-treated compared to raw
chickpea seeds.
In order to understand the developmental changes undergone in seeds during soaking,
the % fresh weight gain profile of alfalfa seeds was determined as shown in Figure 4.2 Seeds
are known to be quiescent and can be stored for months without harm, but once supplied with
water, they become hydrated again and embark on a different stage of activity. This stage
results in outgrowth of the root and later the shoot at the expense of the reserve materials. The
imbibition profile was thus studied in order to determine the kinetics of this new pattern of
development and to understand how HHP possibly interfered with this developmental
process. The imbibition curve was found to exhibit a biphasic pattern of water uptake
characterized by a rapid increase in fresh weight during the first 3 hrs of imbibition (Phase I)
followed by a much slower rate of water uptake lasting for approximately 15 hrs (Phase II).
Dry seeds by virtue of their low water potentials set up a very high water potential gradient as

108
soon as they become in contact with water resulting in imbibition. This helps to explain the
rapid uptake of water shown in Figure 4.2 which occurred at a fairly uniform rate within the
first 3 hrs. Imbibition then slackened off leading to a slower rate of water uptake and
plateaued out until no further fresh weight gain could be observed. This point was coincided
by the onset of sprouting marked by the protrusion of the radicle from the seed, which
occurred 18 hrs after soaking.
As mentioned previously, we observed that an hour of soaking before HHP treatment
was optimal and soaking beyond an hour brought about a gradual decrease in the seeds
viability after pressure treatment. Reconciling the germination rate data shown in Table 4.4
with the imbibition profile of Figure 4.2, we attribute this critical time-dependence to the fact
that imbibing seeds may not swell uniformly throughout their tissues. In pea seeds for
example, it has been shown that the testa is completely wetted at a relatively early stage of
imbibition (Houben, 1966). It is likely that during the first hour of soaking, the testa swells to
the extent of leaving a space between the seed coat and the embryo. This space, probably
water-filled, acts as a protective cushion for the seed embryo during HHP. More prolonged
soaking however may cause water to permeate all the way into the embryo which also swells,
eventually occupying the whole volume inside the testa causing the embryo to stretch the
slightly elastic testa. This has been demonstrated to happen in pea seeds after 4-5 hrs of
soaking (Houben, 1966). Hence controlling the length of soaking is highly critical in order to
ensure complete decontamination of seeds while maintaining a high germination yield.

109
Effect of reduced soaking time and extended pressure holding time at 600 MPa on
inactivation of E. coli O157: H7 and on the germination rate of seeds
Since previous findings showed that the process of soaking seeds for an hour followed
by HHP treatment was able to eliminate pathogens at a slight expense on the germination rate,
we thus investigated whether seeds could be decontaminated by shortening the soaking time
whilst extending the pressure holding time in order to alleviate the structural damage of HHP
on pre-soaked seeds. Results for the inactivation and germination experiments with seeds
soaked and pressure-treated for various time combinations are shown in Table 4.5 and Figure
4.3, respectively. Elimination was consistently achieved when the soaking time was 10 or 15
min with a pressure exposure time of 15 min. In addition, un-inoculated seeds treated under
these conditions still retained their viability to a large extent (Figure 4.3). The process of
soaking seeds for 15 min followed by pressure treatment of 15 min achieved a final
germination yield of 89% which was 4% lower than the control. Differences in seed
germination rate between the control and either treatment on the same day of germination
were not statistically significant (P > 0.05).
Conclusions
Results of this study demonstrate that the application of oscillatory pressurization
produced little enhancement in pressure inactivation of E. coli O157:H7 on alfalfa seeds.
Although the degree of pressure inactivation increased as a function of the number of cycles,
oscillatory HHP treatment could not achieve a > 5 log reduction even after five 2-min cycles
at 600 MPa. This work also provides evidence that pressure treatments in conjunction with
antimicrobials including sanitizers, acidulants or a combination of sanitizer and surfactant

110
were not sufficient to eliminate E. coli O157:H7 on alfalfa seeds. However, when seeds were
soaked in water for 60 min and subsequently treated at 600 MPa for 2 min, elimination of E.
coli O157:H7 occurred at a slight expense on the seed germinability. In addition, it was
demonstrated that seeds could be alternatively processed by a brief soaking of 10 min in water
followed by an extended pressure treatment of 15 min to achieve the same goals with respect
to microbial safety and viability of alfalfa seeds.

111
References Anonymous, 1999. Microbiological safety evaluations and recommendations on sprouted seeds. International Journal of Food Microbiology 52, 123-153. Aleman, G.D., Ting, E.Y., Mordre, S.C., Hawes, A.C.O., Walker, M., Farkas, D.F., Torres, J.A.,1996. Pulsed ultra high pressure treatments for pasteurization of pineapple juice. Journal of Food Science 61, 389-390. Ariefdjohan, M.W., Nelson, P.E., Singh, R.K., Bhunia, A.K., Balasubramaniam, V.M., Singh, N., 2004. Efficacy of high hydrostatic pressure treatment in reducing Escherichia coli O157 and Listeria monocytogenes in alfalfa seeds. Journal of Food Science 69, 117-126. Beuchat, L.R., Scouten, A.J., 2002. Combined effects of water activity, temperature and chemical treatments on the survival of Salmonella and Escherichia coli O157:H7 on alfalfa seeds. Journal of Applied Microbiology 92, 382-395. Blaszczak, W., Doblado, R., Frias, J., Vidal-Valverde, C., Sadowska, J., Fornal, J.,2007. Microstructural and biochemical changes in raw and germinated cowpea seeds upon high-pressure treatment. Food Research International 40, 415-423. Brooks, J.T., Rowe, S.Y., Shillam, P., Heltzel, D.M., Hunter, S.B., Slutsker, L., Hoekstra, R.M., Luby, S.P., 2001. Salmonella Typhimurium infections transmitted by chlorine-pretreated clover sprout seeds. American Journal of Epidemiology 154, 1020-1028. Chen, H. 2007. Use of linear, Weibull, and log-logistic functions to model pressure inactivation of seven foodborne pathogens in milk. Food Microbiology 24, 197-204. Delaquis, P.J., Sholberg, P.L., Stanich, K.,1999. Disinfection of mung bean seed with gaseous acetic acid. Journal of Food Protection 62, 953-957. Earnshaw, R.G., Appleyard, J., Hurst, R.M.,1995. Understanding physical inactivation processes: combined preservation opportunities using heat, ultrasound and pressure. International Journal of Food Microbiology 28, 197-219. Ferguson, D.D., Scheftel, J., Cronquist, A., Smith, K., Woo-Ming, A., Anderson, E., Knutsen, J., De, A.K., Gershman, K., 2005. Temporally distinct Escherichia coli O157 outbreaks associated with alfalfa sprouts linked to a common seed source – Colorado and Minnesota, 2003. Epidemiology and Infection 133, 439-447. Fett, W.F., 2002. Reduction of Escherichia coli O157:H7 and Salmonella spp. on laboratory-inoculated mung bean seed by chlorine treatment. Journal of Food Protection 65, 848-852.

112
Hayakawa, I., Furukawa, S., Midzunaga, A., Horiuchi, H., Nakashima, T., Fujio, Y., Yano, Y., Ishikura, T., Sasaki, K.,1998. Mechanism of inactivation of heat-tolerant spores of Bacillus stearothermophilus IFO 12550 by rapid decompression. Journal of Food Science 63, 371-374. Houben, J., 1966. Imbibition des graines de Pisum Sativum 1ere partie Imbibition par Trempage (Imbibition of seeds of Pisum Sativum 1st stage of imbibition by soaking). Planta 71, 87-97. Jaquette, C.B., Beuchat, L.R., Mahon, B.E.,1996. Efficacy of chlorine and heat treatment in killing Salmonella stanley inoculated onto alfalfa seeds and growth and survival of the pathogen during sprouting and storage. Applied and Environmental Microbiology 62, 2212-2215. Kingsley, D.H., Guan, D., Hoover, D.G., Chen, H., 2006. Inactivation of Hepatitis A Virus by High-Pressure Processing: The Role of Temperature and Pressure Oscillation. Journal of Food Protection 69, 2454-2459. Kingsley, D.H., Chen, H., 2008. Aqueous matrix compositions and pH influence virus inactivation by high pressure processing. Journal of Food Protection 71, 1598-1603. Kumar, M., Hora, R., Kostrzynska, M., Waites, W.M., Warriner, K.,2006. Inactivation of Escherichia coli O157:H7 and Salmonella on mung beans, alfalfa, and other seed types destined for sprout production by using an oxychloro-based sanitizer. Journal of Food Protection 69, 1571-1578. Kurtzweil, P., 1999. Questions keep sprouting about sprouts. FDA Consumer 33, 18-22. Lang, M.M., Ingham, B.H., Ingham, S.C., 2000. Efficacy of novel organic acid and hypochlorite treatments for eliminating Escherichia coli O157:H7 from alfalfa seeds prior to sprouting. International Journal of Food Microbiology 58, 73-82. Neetoo, H., Ye, M., Chen, H., 2008. Potential application of high hydrostatic pressure to eliminate Escherichia coli O157:H7 on alfalfa sprouted seeds. International Journal of Food Microbiology 128, 348-53. Oxen, P., Knorr, D.,1993. Baroprotective effects of high solute concentrations against inactivation of Rhodotorula rubra. LWT Food Science & Technology 26, 220-223. Palou, E., Lopez-Malo, A., Barbosa-Canovas, G.V., Welti-Chanes, J., Swanson, B.G., 1997. Effect of water activity on high hydrostatic pressure inhibition of Zygosaccharomyces bailii. Letters in Applied Microbiology 24, 417-420

113
Palou, E., Lopez-Malo, A., Barbosa-Canovas, G.V., Welti-Chanes, J., Davidson, P.M., Swanson, B.G.,1998. Effect of oscillatory high hydrostatic pressure treatments on Byssochlamys nivea ascospores suspended in fruit juice concentrates. Letters in Applied Microbiology 27, 375-378. Palou, E., Lopez-Malo, A., Barbosa-Canovas, G.V., Welti-Chanes, J., Swanson, B.G.,1998. Oscillatory high hydrostatic pressure inactivation of Zygosaccharomyces bailii. Journal of Food Protection 61, 1213-1215. Piernas, V., Guiraud, J.P.,1997. Disinfection of rice seeds prior to sprouting. Journal of Food Science 62, 611-615. Proctor, M.E., Hamacher, M., Tortorello, M.L., Archer, J.R., Davis, J.P., 2001. Multistate outbreak of Salmonella serovar Muenchen infections associated with alfalfa sprouts grown from seeds pretreated with calcium hypochlorite. Journal Clinical Microbiology 39, 3461-3465. Taormina, P.J., Beuchat, L.R., 1999. Comparison of chemical treatments to eliminate enterohemorrhagic Escherichia coli O157:H7 on alfalfa seeds. Journal of Food Protection 62, 318-324. Weissinger, W.R., Beuchat, L.R., 2000. Comparison of aqueous chemical treatments to eliminate Salmonella on alfalfa seeds. Journal of Food Protection 63, 1475-1482. Wuytack, E.Y., Michiels, C.W., Meersseman, K., Diels, A.M., 2003. Decontamination of seeds for seed sprout production by high hydrostatic pressure. Journal of Food Protection 66, 918-923.

114
Figure 4.1.
Figure 4.2.

115
Figure 4.3.

116
Table 4.1. Effect of HHP (600 MPa for 2 min at 20°C) in conjunction with antibacterial agents on inactivation of E. coli O157:H7 on alfalfa seeds inoculated at a level of 2.0 x 105 CFU/g
Numbers separated by a “/” represent the number of samples testing positive after enrichment out of a total of three trials. Table 4.2. Effect of extended HHP holding times (600 MPa and 20°C) in conjunction with antibacterial agents on inactivation of E. coli O157:H7 on alfalfa seeds inoculated at a level of 1.2 x 105
CFU/g Pressure holding time (min) Fluid Concentration 5 10 15 Water 0 3/3 3/3 3/3 Ca(OCL)2 2000 ppm 3/3 3/3 3/3 Ca(OCL)2 20000 ppm 3/3 3/3 3/3 Ca (OH)2 1% 3/3 3/3 3/3 Ca (OH)2 1% + 1% Tween 80 3/3 3/3 3/3 Lactic acid 1% 3/3 3/3 3/3 Lactic acid 2% 3/3 3/3 3/3 Lactic acid 5% 3/3 3/3 3/3 Numbers separated by a “/” represent the number of samples testing positive after enrichment out of a total of three trials.
Soaking time (min) Fluid Concentration 0 5 10 15 Water 0 3/3 3/3 3/3 3/3 Ca(OCL)2 200 ppm 3/3 3/3 3/3 3/3 Ca(OCL)2 1000 ppm 3/3 3/3 3/3 3/3 Ca(OCL)2 2000 ppm 3/3 3/3 3/3 3/3 Ca(OCL)2 20, 000 ppm 3/3 3/3 3/3 3/3 Ca (OH)2 1% 3/3 3/3 3/3 3/3 Ca (OH)2 1% + 1% Tween 80 3/3 3/3 3/3 3/3 Lactic acid 1% 3/3 3/3 3/3 3/3 Lactic acid 2% 3/3 3/3 3/3 3/3 Lactic acid 5% 3/3 3/3 3/3 3/3 Sodium acid sulfate 0.05% 3/3 3/3 3/3 3/3 Sodium acid sulfate 0.10% 3/3 3/3 3/3 3/3 Sodium acid sulfate 0.20% 3/3 3/3 3/3 3/3

117
Table 4.3. Effect of soaking time prior to treatment at 600 MPa for 2 min at 20°C on inactivation of E. coli O157:H7 on alfalfa seeds inoculated at a level of 2.5 x 105 CFU/g Water Soaking time prior to HHP (min) (ml) 0 30 60 90 120 180 240 300
3 2.7±0.8a 1.5±0.1b 1.1±0.4b 1.4±0.1b 1/3 1/3 0/3 0/3 20 2.7±0.8 1/3 0/3 0/3 0/3 0/3 0/3 0/3
Data are the means of log survivors (CFU/g) ± one standard deviation. Numbers separated by a “/” represent the number of samples testing positive after enrichment out of a total of three trials (The counts were below the detection limit by direct plating (0.8 log CFU/g) in the three trials). Data in the same row followed by the same letter are not significantly different (P > 0.05).
Table 4.4. Effect of soaking time prior to treatment at 600 MPa for 2 min at 20°C on the germination rate of alfalfa seeds Soaking time prior to HHP (min) Days of germination control 0 30 60 90 120 180
3 87±4a 80±5ac 75±6ad 80±6ae 71±7a 60±12bcde 40± 8b 4 89±3a 87±6a 79±7a 85±6a 75±8a 67±11ac 47±10bc 5 91±4a 91±7a 81±7a 88±6a 78±8a 73±10ac 52±11bc 6 93±3a 93±7a 82±7a 89±6a 80±9a 76±11ac 55±1bc 7 95±3a 95±6a 84±7a 90±6a 81±9a 79±11ac 58±13bc 8 95±3a 95±6a 86±7ac 91±5a 84±9ad 82±13ae 62±13bcde
Data are the means of % germination ± one standard deviation. Data for the same day of germination followed by the same letter are not significantly different (P > 0.05). Table 4.5. Effect of reduced soaking time and extended pressure holding time on inactivation of E. coli O157: H7 on alfalfa seeds inoculated at a level of 2.5 x 105 CFU/g Holding time at Soaking time prior to HHP (min) 600 MPa (min) 10 15
5 3/3 2/3 10 3/3 3/3 15 0/3 0/3
Numbers represent the number of samples testing positive after enrichment out of a total of three trials. The counts were below the detection limit by direct plating (0.8 log CFU/g) throughout.

118
Chapter 5
Conditions for elimination of Escherichia coli O157:H7 on alfalfa seeds through a
combination of high hydrostatic pressure and mild heat
Published in Appl. Environ. Microbiol. 2009. 75, 1901-1907

119
Abstract
Escherichia coli O157:H7 have been associated with contaminated seed sprout
outbreaks. The majority of these outbreaks have been traced to sprout seeds contaminated
with low levels of pathogens. Sanitizing sprout seeds presents a unique challenge in the arena
of produce safety in that even a low residual pathogen population remaining on contaminated
seed after treatments appears capable of growing to very high levels during sprouting. In this
study, the effectiveness of high pressure treatment in combination with low and elevated
temperatures was assessed for its ability to eliminate E. coli O157:H7 on artificially
contaminated alfalfa seeds. Inoculated seed samples were treated at 600 MPa for 2 min at 4,
20, 25, 30, 35, 40, 45 and 50°C. The pressure-sensitivity of the pathogenic bacteria was
strongly dependent on the treatment temperature. At 40°C, the process was adequate in
eliminating a 5 log population on the seeds with no adverse effect on the seed viability.
Treatments carried out at reduced pressure levels and/or extended treatment time, 550 MPa
for 2 min at 40°C, 300 MPa for 2 min at 50°C, and 400 MPa for 5 min at 45°C were equally
lethal to the pathogen. When all the three treatments were compared in terms of their impact
on the seed viability, the process of 550 MPa for 2 min at 40°C was the most desirable
achieving final germination percentages and sprout sizes statistically similar to control
untreated seeds (P > 0.05).

120
Introduction
In many countries worldwide, including the United States, the consumption of seed sprouts
has increased in recent decades with the advent of neutraceuticals, phytochemicals and the
shift of consumer preference towards health foods (Gabriel et al., 2007; Pandrangi et al.,
2003). Alfalfa sprouts are one of the most common sprouts consumed in the United States
due to availability and nutritional value (Fett et al., 2005; Meyerowitz, 1999). However, the
sprouting process makes the commodity susceptible to microbial contamination and growth,
thereby compromising the safety and quality of the sprouts (Gabriel, 2005).
Recently, alfalfa and other types of sprouts that are often consumed raw have been
designated as a special food safety problem by the National Advisory Committee on
Microbiological Criteria for Foods because of the propensity of human pathogens such as
Escherichia coli O157:H7 to rapidly multiply on sprouts during sprouting and because of lack
of a post-harvest lethal step in the processing of sprouts (NACMCF, 1999). Since 1995 in the
United States, there have been two outbreaks caused by E. coli O157 (O157:H7 and O157:
NM) due to the consumption of contaminated alfalfa and clover sprouts (Breuer et al., 2001;
Ferguson et al., 2003; MMWR, 1997, Mohle-Boetani et al., 2001). In most sprout-associated
outbreaks, the initial source of contamination was thought to be the seeds themselves on the
basis of direct isolation and/or epidemiological evidence (Fett et al., 2005).
Many studies have thus been performed to determine the effectiveness of a wide range
of treatments for reducing the levels of pathogenic E. coli O157:H7 on seeds and sprouts

121
(Bari et al., 2008). Decontamination by soaking the seeds in different chemical solutions has
been thoroughly investigated. Chemicals used include chlorine, organic acids, hydrogen
peroxide and ethanol and these have been reported to have varying degrees of efficacy (Bari
et al., 2003; Beuchat et al., 2001; Jaquette et al., 1996). Very often, the bacterial cells may
reside in the seed crevices or between the seed coat and cotyledon (Charkowski et al., 2001),
providing protection from chemical sanitation. Sprouting conditions may provide a suitable
temperature and moisture conditions for the growth of E. coli O157:H7 and Salmonella.
These conditions together with nutrients released by sprouting seeds, help low levels of
attached pathogens reach dangerous levels on the finished sprouts (Stewart et al., 2001).
Conventional thermal processing has the potential to ensure food safety and lead to an
extended shelf-life. However, it often leads to detrimental changes in the sensory and
nutritional qualities of the product (Munoz et al., 2007). With non-thermal processing
technologies such as high hydrostatic pressure (HHP) technology, more fresh-like products
can be obtained. HHP treatment is advantageous because of its ability to inactivate a number
of enzymes and microorganisms contained in foods without significantly altering their
sensory and nutritional properties (Arroyo et al., 1999).
The application of HHP alone to enhance the safety or quality of seeds or sprouts have
been studied previously (Ariefdjohan et al., 2004; , Munoz et al., 2006; Neetoo et al., 2008;
Penas et al., 2008; Wuytack et al., 2003) with reported varying degrees of efficacy. Research
in our laboratory demonstrated that the application of pressure at a level of 650 MPa for 15
minutes at 20°C was adequate in eliminating E. coli O157:H7 from alfalfa seeds (Neetoo et
al., 2008). The aim of this study was to develop a high pressure process to achieve reduction

122
of a 5 log initial population of E. coli O157:H7 on alfalfa seeds to undetectable level (cells
were not detected using an enrichment method) and subsequently assess the effects of the
process on the viability retention of the seeds. To simplify sentence constructions, the term
“elimination” was used in this paper interchangeably with “reduction of the bacterial
population to undetectable level”.
Materials and Methods
Bacterial strains and preparation of inoculum
E. coli O157:H7 strains 1730, 250, 251, J58 and Cider were used. The source of the strains
was provided in the paper by Neetoo et al. (2008). The cells of E. coli O157:H7 were adapted
to grow in tryptic soy broth plus 0.6% yeast extract (Difco Laboratories, Sparks, MD, USA)
supplemented with 50 µg/ml of nalidixic acid (Fisher Scientific, Hampton, NH, USA)
(TSBYE-N). Individual cultures were grown in TSBYE-N overnight at 35°C. Cultures were
then transferred (one loopful) into 10 ml of fresh TSBYE-N and incubated at 35°C for 24 h.
Equal volumes of individual cultures were mixed to form a five-strain cocktail of E. coli
O157:H7.
Inoculation of seeds
The cocktail (10 ml) was mixed with 100 ml of sterile 0.1% peptone water (Fisher). Alfalfa
seeds (Medicago sativa) (100 g), obtained from International Specialty Supply (Cookeville,
Tenn., USA), were added to the cell suspension and gently stirred for 5 min. The seeds were
separated from the cell suspension by pouring the mixture over a double layer of cheesecloth
supported by a wire screen and dried inside a bio-safety hood at room temperature (21 ± 1°C)
for 24 h. Dried seeds (aw ~ 0.622) with an approximate inoculation level of 105 CFU/g of

123
E. coli O157:H7 were placed in sterile pouches and stored at 4°C.
High Pressure Treatment
Two g of inoculated seeds and 3 ml of sterile DI water were placed in a 3-mil thick pouch
(Nylon/Polyethylene, Koch Supplies, Kansas City, MO). To avoid leakage during pressure-
treatment, each sample pouch was placed in a larger pouch of an 8-mil thick PVC plastic
(McMaster-Carr, Elmhurst, IL) and heat-sealed. HHP treatment of samples was carried out
using a high-pressure unit with temperature control (Model Avure PT-1, Avure Technologies,
Kent, WA). The experiments were conducted at temperatures ranging from 4°C to 50°C
(initial seed sample temperature prior to pressure treatment) using water as a hydrostatic
medium at a pressure level of 600 MPa and a treatment time of 2 min. Samples were
submerged in the water bath surrounding the pressure cell for 10 min for samples to
equilibrate to the water bath temperature before pressurization. The pressure-come-up rate
was approximately 22 MPa/s. The pressure-release was almost immediate (< 4s).
Pressurization time reported in this study did not include the pressure come-up or release
times.
Microbiological analysis
Pouches containing treated seeds were cut open aseptically. The sample was transferred into a
stomacher bag to which 8 ml of sterile 0.1% peptone water was added and subsequently
stomached for 2 min at 260 rpm (Seward 400 Stomacher, Seward Medical Co., London,
U.K.). The seed slurry was serially diluted in sterile 0.1% peptone and surface-plated in
duplicate on tryptic soy agar with 0.6% yeast extract (Difco Laboratories, Sparks, MD, USA)
supplemented with nalidixic acid to a final concentration of 50 µg/ml (TSAYE-N). TSAYE-N

124
plates were incubated for 3 days at 35ºC. Presumptive colonies of E. coli O157:H7 formed on
the plates were enumerated. Occasionally, colonies were confirmed to be E. coli O157:H7
using either a BAX® system PCR assay for screening E. coli O157:H7 (Qualicon-DuPont,
Wilmington, DE, USA) or Rapid E. coli O157:H7 Test Methods (Strategic Diagnostics Inc.,
Newark, DE, USA). Seed slurry samples were also directly enriched in 90 ml of TSBYE-N
and incubated for 48 h at 35ºC to allow resuscitation of sub-lethally injured cells. Samples
were streaked onto Sorbitol MacConkey agar (Difco Laboratories, Sparks, MD, U.S.A.) plates
supplemented with 50 µg/ml of nalidixic acid. After 24 h incubation, presence of growth
exhibiting morphological and biochemical characteristics typical of E. coli O157:H7 were
determined by visually inspecting the plates.
Determination of % germination of pressure-treated alfalfa seeds
To determine the effect of pressure-treatment on the seeds germination potential, 2 g of un-
inoculated seeds were introduced into a pouch to which 3 ml of DI-water was added. The
samples were treated at 600 MPa for 2 min at 40°C. Pressure-treated and untreated seeds
(control) were soaked in DI-water for 3 hrs. One hundred seeds were drawn from the soaked
seeds and spread evenly on pieces of wet paper towels on a plastic rack, which in turn was
placed into a water-filled bucket to provide a moist environment for the seeds. The water level
was maintained below the seeds’ level. The bucket was kept at room temperature for 8 days
(suggested by the seeds provider) and misted daily. The bucket was covered loosely with a
piece of plastic film to allow exchange of air between the inside and outside of the bucket.
Sprouted seeds characterized by the emergence of the root tip (radicle) were enumerated 3 to

125
8 days after setting up the germination system and the determinations recorded as %
Germination.
Application of mild heat and reduced pressure levels and/or extended treatment time on
the inactivation of E. coli O157:H7
Two g of inoculated seeds were mixed with 3 ml of sterile DI-water, packaged and treated at
250-550 MPa for 2 min at 40, 45 and 50°C or at 300-500 MPa for 5 min at 40 and 45°C.
Immediately after pressure treatment, samples were cooled in an ice-water mixture. Samples
were then microbiologically assayed as described previously and enriched for the detection of
survivors. In addition, inoculated samples were immersed for 10 min in the water bath at
50°C followed by a 3 min immersion at 61°C to determine the effect of temperature alone on
the inactivation of E. coli O157:H7.
Assessment of germination rate and sprout length of HHP-treated seeds of different ages
Two g of un-inoculated seeds from either a freshly received or an old batch of alfalfa seeds
(nine months storage at room temperature after receipt) were mixed with 3 ml of DI-water,
packaged and treated at 300 MPa for 2 min at 50°C, 400 MPa for 5 min at 45°C and 550 MPa
for 2 min at 40°C. One hundred seeds were drawn from the pressure-treated and untreated
(control) seeds and assayed for germinability as described above. Fifty seeds were also drawn
from the treated and untreated samples and allowed to germinate for 8 days. The sprout
lengths of germinated alfalfa seeds were determined with a digital vernier caliper.
Validation of the selected optimum HHP process
Two g of inoculated seeds from a freshly received or an old batch of alfalfa seeds (12 months
storage at room temperature after receipt) were mixed with 3 ml of sterile DI-water, packaged

126
and treated at 550 MPa for 2 min at 40°C. Immediately after pressure treatment, samples were
cooled in an ice-water mixture. Samples were then microbiologically assayed as described
previously and enriched for the detection of survivors. In addition, the same procedure was
repeated for un-inoculated seeds from the same batch of fresh and old seeds. One hundred
seeds were drawn from the pressure-treated (aged and fresh) seeds and their untreated
counterparts and assayed for germinability as described above. To evaluate the reliability of
the process, a total of ten trials were carried out for both pathogen inactivation and
germination tests.
Statistical analysis
All experiments were replicated three times with the exception of the validation experiment
which was replicated ten times. Where appropriate, statistical analyses were conducted using
Minitab® Release 15 (Minitab Inc., University Park, PA, USA). One-way analysis of variance
(ANOVA) and Tukey’s one-way multiple comparisons were used to determine differences in
the populations of E. coli O157:H7 recovered on treated alfalfa seeds as well as differences in
the germination rate of seeds. Significant differences were considered at the 95% confidence
level (P < 0.05).
Results
Effect of temperature on pressure inactivation of E. coli O157:H7 on alfalfa seeds and
resulting seed viability
Results for the pressure inactivation of E. coli O157:H7 on alfalfa seeds at different initial
sample temperatures are shown in Figure 5.1. E. coli O157:H7 responded quite similarly to a
pressure level of 600 MPa at initial sample temperatures of 4 and 25°C achieving 2.4-2.8 log

127
CFU/g reduction. When the initial sample temperature was increased to ≥ 30°C, the pressure
treatments resulted in ≥ 5 log reduction of the pathogen although survivors were still detected
post-enrichment for treatments at 30 and 35°C. Survivors were not detected using the
enrichment method when the temperature was raised to ≥ 40°C.
Since a treatment of 600 MPa for 2 min at 40°C was adequate in eliminating a > 5 log
initial population of E. coli O157:H7 on the seeds, seeds pressure-treated under this condition
were assayed for germination. Results are shown in Fig. 5.2. Overall the pressure treatment
did not bring about any statistically significant difference between the control and pressure-
treated seeds (P > 0.05) after eight days of sprouting.
Effect of the combined application of mild heat and reduced pressure levels and/or
extended treatment time on the inactivation of E. coli O157:H7
Since a combination of high pressure of 600 MPa and elevated temperature of 40°C was able
to eliminate the pathogen on the seeds, further experiments were conducted to investigate the
application of lower pressure levels in combination with mild heat to achieve an equivalent
kill. Results for this experiment are summarized in Table 5.1. At 40°C and 45°C, the lowest
pressure that could achieve consistent reduction of the pathogens to undetectable level during
a 2-min treatment was found to be 550 MPa. By increasing the initial sample temperature to
50°C, a pressure treatment at a magnitude as low as 300 MPa successfully eliminated a ~5 log
population of E. coli O157:H7 on the seeds.
Since the findings showed that a 2-min application of high pressure at a level of 350-
500 MPa at 40 and 45°C resulted in > 5 log inactivation, the pressure treatment time was

128
extended to 5 min to investigate whether a lower pressure level could result in complete kill.
A treatment time of 5 min was chosen to make the process economically feasible. Results for
the 5-min experiment also displayed in Table 5.1 show that the treatments of 300-500 MPa at
40°C resulted in > 5 log reductions, but survivors were still detected at a low frequency.
However, reduction of the pathogens to undetectable levels was consistently achieved (across
three replicates) when a pressure magnitude ranging from 400 MPa to 500 MPa was applied
at 45°C for 5 min. To determine the effect of heat alone in the absence of HHP, inoculated
seed samples were immersed for 10 min in the water bath at 50°C and 3 min at 61°C (to
represent a worst case scenario of adiabatic heat during pressure application of 300 MPa for 2
min at 50°C). It was found that E. coli O157:H7 underwent > 5 log inactivation although
survivors could still be detected post-enrichment (results not shown).
Effect of selected high pressure treatments on the germination rate and sprout lengths of
fresh and aged seeds
Figure 5.3 (A) and (B) compares the germination rates of pressure-treated seeds relative to
untreated seeds for fresh and aged seeds respectively. Untreated fresh seeds had a higher final
germination percentage than their aged counterparts. When fresh seeds were pressure-treated
under the three different conditions, their germination rates underwent a maximum reduction
of 5% relative to the control although the difference was not statistically significant (P >
0.05). The average sprout lengths for fresh and aged seeds subjected to the various pressure-
treatments are shown in Figure 5.4. As far as sprout length is concerned, the treatment of 550
MPa for 2 min at 40°C was the most promising as it had the least impact on sprout growth (P
> 0.05). The pressure-treatment conducted at 50°C, on the other hand, was the least desirable,

129
bringing about the most noticeable decrease in sprout length although the difference was not
significant statistically (P > 0.05). Similar to fresh seeds, the final germination rates of
pressure-treated aged seeds determined after 8 days was found to be reduced by a maximum
of 5% although the difference was not significant statistically (P > 0.05). Aged seeds also
exhibited a decrease in sprout length when pressurized under the different conditions and the
treatment of 550 MPa for 2 min at 40°C was again the most preferable.
Validation of the selected optimum HHP process
It was found that in either case of fresh or aged seeds inoculated with a 5 log of E. coli
O157:H7, of the treatment of 550 MPa for 2 min at 40°C was able to consistently reduce the
populations to undetectable level in all 10 trials conducted. With respect to the seed viability,
ten samples each of un-inoculated fresh and aged seeds were pressure-treated and assayed for
germination. It was found that the average percentage sprouted seeds determined after 3 days
of germination were ~ 80% and ~ 92% for pressure-treated old and fresh seeds respectively
while their untreated counterparts achieved an average germination of ~ 87 and ~ 97%
respectively. After eight days of germination, the final % germination reached > 86 and >
97% for treated old and fresh seeds respectively, while their untreated counterparts reached an
average of 91 and 100% respectively.
Discussion
Effect of temperature on pressure inactivation of E. coli O157:H7 on alfalfa seeds and
resulting seed viability
The results of Fig. 5.1 show that the pressure sensitivity of E. coli O157:H7 had a strong
dependence on the treatment temperature, exhibiting the highest pressure resistance at

130
temperatures ranging from 4 to 25°C and becoming more sensitive the higher the temperature
of the treatment. It is well documented that the temperature of food during pressurization
plays a significant role in the inactivation of bacteria and viruses. In particular, elevated
temperatures above 30°C has been shown to enhance the pressure inactivation of bacteria as
reported for Listeria monocytogenes (Chen, 2007), Staphylococcus aureus (Chen 2006),
Escherichia coli (Ponce et al., 1998), Salmonella enterica serovar Enteritidis (Ponce et al.,
1994), Vibrio vulnificus (Kural and Chen, 2008) and Vibrio parahaemolyticus (Kural et al.,
2008). Studies conducted by Chen et al. (2005) and Kingsley et al. (2006) demonstrated that
temperatures > 30°C also increased the pressure inactivation of Feline calicivirus and hepatitis
A virus (HAV) respectively. However, contrary to other authors (Carlez et al., 1993; Chen,
2007; Chen, 2007; Kural and Chen, 2008; Kural et al., 2008), we did not observe any
enhancement in pressure inactivation at low temperatures (i.e < 20°C). In fact, E. coli
O157:H7 exhibited the highest baro-resistance at the lowest temperature investigated i.e at
4°C. Similarly, Ponce et al. (1999) and Kingsley et al. (2006) showed that the respective
inactivation of Salmonella Enteritidis and HAV was lower at lower temperatures.
Un-inoculated seed samples treated at 600 MPa for 2 min at 40°C did not undergo any
significant difference in germination rates relative to untreated seeds after eight days of
sprouting (P > 0.05). When the adiabatic compression heating component of the pressure
treatment was taken into consideration, the temperature of the samples increased to a
maximum of 59.2°C during pressure treatment. Bari et al. (2008) showed that the use of very
high temperature for short times (hot water treatment at 90°C for 90 s followed by dipping in

131
chilled water for 30 s) resulted in the elimination of Salmonella and E. coli O157:H7 and the
process did not affect the germination yield of mung bean seeds to any significant extent.
Weiss and Hammes (2005) indicated that the treatment of alfalfa and radish seeds with hot
water could be an effective seed sanitizing step achieving > 95% germination. They
demonstrated that a treatment of seed inoculated with Salmonella sp. or E. coli O157:H7 for
0.5-8 min at 53-64°C reduced all pathogens by more than 5 log CFU/g although no evidence
of elimination was provided. Hence, studies conducted by other authors have pointed to the
possible application of brief high-temperature treatment to inactivate seed-borne pathogenic
microorganisms with minimal impact on the seed viability. However, the drawback associated
with high temperature treatments is the reported lack of consistency of the effect of the
treatments on the seed viability (Jaquette et al., 1996) and on the safety of sprouts. Under
commercial practice, the ability of hot water treatments to ensure consistent elimination of
bacterial human pathogens from alfalfa seed was put into question by a recent multi-state
outbreak of salmonellosis due to contaminated alfalfa sprouts grown from seed treated with
hot water followed by a soak in low levels (2000 ppm) of chlorine (Winthrop, 2003).
Neetoo et al. (2008) recently demonstrated that pressure-treatment at 600 MPa for 20
min at 20°C was inadequate in completely eliminating E. coli O157:H7 from seeds. In the
current study, the combined application of pressure and mild temperatures to enhance the
safety of alfalfa seeds was investigated. By increasing the initial sample temperature to 40°C,
the HHP process at 600 MPa proved to be an effective intervention technology to enhance the
safety of alfalfa seeds. Both Penas et al. (2008) and Ariefdjohan et al. (2004) also
investigated the application of high pressure on alfalfa seeds to improve the safety of the

132
resultant sprouts. Penas et al. (2008) found that seeds pre-soaked in water and subsequently
pressure-treated at 400 MPa and 40°C in the absence of water exhibited a significantly lower
germination rate (< 30%) than untreated alfalfa seeds. On the other hand, Ariefdjohan et al.
(2004) found that pressurizing dry (non pre-soaked) alfalfa seeds at 40°C (275–575MPa for 2
min or 475 MPa for 2–8 min) took longer to germinate, achieving final germination of up to
34%, while 95% of the control seed germinated. Unlike findings reported by those authors,
our data show that alfalfa seeds pressure-treated in the presence of water at the same
temperature of 40°C and at an even higher pressure magnitude of 600 MPa still achieved a
final germination percentage comparable to the control (P > 0.05). It is most likely because
the other authors pressure-treated pre-soaked (Penas et al., 2008) or dry (Ariefdjohan et al.,
2004) seeds in the absence of water while in our study, seeds were pressure-treated whilst
immersed in water. In order to demonstrate the adverse effect of pressure treatment of pre-
soaked seeds in the absence of water, we immersed alfalfa seeds in de-ionized water for 3
hours and then pressure-treated them un-immersed in water at 600 MPa for 2 min at 40°C.
The average final germination percentage of pressure-treated seeds after eight days of
germination was ~19%. On the other hand, seeds pressure-treated using the process we
developed consisting of HHP treatment of seeds immersed in water without prior soaking,
resulted in a significantly higher final germination percentage of 96%. It is possible that the
presence of immersion water surrounding the seeds acts as a cushion protecting the seeds
against any physical and structural damage during pressure-treatment.

133
Effect of the combined application of mild heat and reduced pressure levels and/or
extended treatment time on the inactivation of E. coli O157:H7
The experimental data of Table 5.1 displays a slight degree of inter-experimental variability
as evidenced by the positive enrichment results for the two treatments of 350 MPa and 45°C,
1/3 for the 2-min treatment and 2/3 for the 5-min treatment. It would be expected that
increasing the treatment time would increase the lethal effect on microorganisms. However, it
has been widely reported that pressure inactivation of microorganisms is not always first order
and the tailing effect of survival curves has been found in many species including E. coli
O157:H7 (Chen, 2007). The occurrence of tails is usually attributed to the presence of a more
resistant subpopulation of cells. Therefore a small increase in treatment time may not
necessarily be able to enhance the inactivation effect on these resistant cells. Chance of
bacterial elimination might also depend on the samples being treated which might have slight
variations with respect to cell age or genetic constitution of the inoculum.
Results of Table 5.1 demonstrate that the pressure level of the treatment could be
substantially reduced by increasing the sample temperature to achieve an equivalent lethality.
Since the capital costs of the high-pressure equipment increase exponentially with operating
pressures and process costs are related to operating pressure (Chen, 2007), it is economically
beneficial to use lower levels of pressure in combination with other hurdles such as elevated
temperatures to inactivate E. coli O157:H7 on alfalfa seeds. Munoz et al. (2007) showed that
the inactivation of microorganisms in orange juice and vegetable soup increased when the
temperature was increased from 20°C to 60°C demonstrating that a treatment at 60°C brought
about the greatest reductions during the application of high pressure. Hence, it can be

134
concluded that elevated temperature can reduce the pressure magnitude needed to achieve a
target level of pathogen inactivation. The manipulation of the process parameters can thus
greatly influence the outcome and economics of a high-pressure process.
In addition, Table 5.1. shows that the combinations that were effective in eliminating
an initial population of ~ 5 log of E. coli O157:H7 included treatments at a pressure ≥ 550
MPa for 2 min at 40°C (maximum temperature reached due to adiabatic heating was 57.6°C at
550 MPa), treatments at a pressure ≥ 400 MPa for 5 min at 45°C (maximum temperature
reached due to adiabatic heating was 58.2°C at 400 MPa), and treatments at a pressure ≥ 300
MPa for 2 min at 50°C (maximum temperature reached due to adiabatic heating was 60.2°C at
300 MPa). Temperature changes are known to occur during compression and pressure-release
as a result of adiabatic heating and cooling (Ting et al., 2002). This adiabatic temperature
change is predictable; all compressible substances will change temperature during physical
compression as a result of this unavoidable thermodynamic effect (Ting et al., 2002). The
magnitude of this change depends mainly on the compressibility of the substance, its specific
heat and the initial sample temperature. Since the early work of Bridgman (1912), the
adiabatic compression behavior of water has been well characterized. At the pressures
typically encountered during HHP (400 – 1000 MPa), under adiabatic conditions near room
temperature, water (the hydrostatic medium used) typically changes by about 3°C for every
100 MPa pressure change. Since the seeds were immersed in water during pressure-treatment,
the adiabatic heating compression factors of pressure-treated seeds were very similar to those
of water. Temperature increases during pressure treatment of the samples were 2.8, 2.9, 3.2,

135
and 3.4°C/100 MPa at 20, 30, 40, and 50°C, respectively. For samples pressure-treated at the
highest starting temperature of 50°C (the harshest treatment) to a final come-up pressure of
300 MPa, the sample temperature rose to a maximum of 60.2°C. Inoculated seed samples
subjected to a control thermal treatment at 61°C still harbored E. coli O157:H7 survivors.
This finding therefore indicates that the thermal effect during high pressure processing was
inadequate on its own in eliminating E. coli O157:H7 pointing to the critical contribution of
pressure-treatment in achieving a complete kill.
Effect of selected high pressure treatments on the germination rate and sprout lengths of
fresh and aged seeds
Since the germinability of alfalfa seeds is known to decrease with the age of the seeds
(Griggs, 2004, Nienhuis and Baltjes, 1985), it was deemed worthwhile to determine the effect
of the pressure treatments on the germination characteristics of treated seeds of different ages.
The germination percentages and the average sprout lengths of fresh and aged seeds pressure-
treated at 550 MPa for 2 min at 40°C, 400 MPa for 5 min at 45°C or 300 MPa for 2 min at
50°C were affected to a variable extent depending on the treatment conditions (Figures 5.3
and 5.4). The treatment at 550 MPa at 40°C was the most promising as it had no significant
adverse effect on the seeds’ viability (P > 0.05). When the seeds were observed under the
light microscope, no differences could be discerned between untreated seeds and seeds
pressurized at 550 MPa at 40°C (results not shown). This contrasts with observations made by
Ariefdjohan et al. (2004) who pressure-treated seeds at 275-575 MPa for 2 min at 40°C and
observed structural damage to the seeds with a concomitant decrease in viability. We attribute

136
this difference to the fact that our seeds were pressurized whilst submerged in water thus
alleviating the seed damage and retaining the germination capability. In the light of these
results, we recommend a pressure treatment of 550 MPa for 2 min at 40°C on alfalfa seeds
since this condition has the ability to eliminate E. coli O157:H7 on seeds whilst having the
least impact on the germination rate and sprout lengths of seeds regardless of the age of the
seeds.
Validation of the selected optimum HHP process
The pressure treatment developed in this study carried out at 550 MPa for 2 min at
40°C was able to consistently reduce a 5-log pathogen load on aged and fresh seeds to
undetectable level throughout all trials with minimal impact on seed viability. It should be
pointed out that small samples of seeds (2 g) were used in this study due to the limitation of
our pressure unit. A scale-up study using a much larger quantity of seeds would be needed
before commercialization of this process. Although it is anticipated that a large-scale pressure
unit would differ with respect to pressure come-up-time and temperature profile of seeds
during pressurization, these differences are not expected to affect the inactivation efficacy and
seed viability retention significantly. In addition, we recommend testing the robustness of the
developed pressure process by conducting study on seeds from different sources.
Conclusion
The application of pressure treatment at optimum temperatures on alfalfa seeds is
highly desirable since results in this study demonstrated that temperature plays a significant
role in pressure inactivation of E. coli O157:H7. Although high pressure processing is more
expensive than the currently available methods for seed decontamination of seeds, HHP

137
appears to be highly effective against E. coli O157:H7 with minimal adverse impact on seed
viability. Since alfalfa seeds are very small (~2 mm long), it is anticipated that a large number
of seeds could be processed at one time thus helping to achieve a reasonably high throughput.
In addition, through a combination of high pressure processing with additional hurdles such as
mild heat and optimization of the various processing parameters (pressure, time and
temperature), the pressure levels needed to achieve a desired target level of bacterial
inactivation can be substantially reduced. Therefore HHP could be feasibly used for
enhancing the safety of sprouts as these niche products are becoming increasingly popular
natural foods.
Acknowledgment We thank Dr. Rolf Joerger (University of Delaware) for his technical support.

138
References
1. Ariefdjohan, M. W., P. E. Nelson, R. K. Singh, A. K. Bhunia, V. M. Balasubramaniam, and N. Singh. 2004. Efficacy of high hydrostatic pressure treatment in reducing Escherichia coli O157 and Listeria monocytogenes in alfalfa seeds. J. Food Sci. 69:117-126. 2. Arroyo, G., P. D. Sanz, and G. Prestamo. 1999. Response to high-pressure, low-temperature treatment in vegetables: determination of survival rates of microbial populations using flow cytometry and detection of peroxidase activity using confocal microscopy. J. Appl. Microbiol. 86:544-556. 3. Bari, M. L., E. Nazuka, Y. Sabina, S. Todoriki, and K. Isshiki. 2003. Chemical and irradiation treatments for killing Escherichia coli O157:H7 on alfalfa, radish, and mung bean seeds. J. Food Prot. 66:767-774. 4. Bari, M. L., Y. Inatsu, S. Isobe, and S. Kawamoto. 2008. Hot water treatments to inactivate Escherichia coli O157:H7 and Salmonella in mung bean seeds. J. Food Prot. 71:830-834. 5. Beuchat, L. R., T. E. Ward, and C. A. Pettigrew. 2001. Comparison of chlorine and a prototype produce wash product for effectiveness in killing Salmonella and Escherichia coli O157:H7 on alfalfa seeds. J. Food Prot. 64:152-158. 6. Breuer, T., D. H. Benkel, R. L. Shapiro, W. N. Hall, M. M. Winnett, M. J. Linn, J. Neimann, T. J. Barrett, S. Dietrich, F. P. Downes, D. M. Toney, J. L. Pearson, H. Rolka, L. Slutsker, and P. M. Griffin. 2001. A multistate outbreak of Escherichia coli O157:H7 infections linked to alfalfa sprouts grown from contaminated seeds. Emerg. Infect. Dis. 7:977-982. 7. Bridgman, P.W. 1912. Water under pressure. Proc. Am. Acad. Arts Sci. 48:450-558. 8. Carlez, A., J. P. Rosec, N. Richard, and J. C. Cheftel. 1993. High pressure inactivation of Citrobacter freundii, Pseudomonas fluorescens and Listeria innocua in inoculated minced beef muscle. Food Sci. Technol. Leb. 26:357-363. 9. Charkowski, A. O., C. Z. Sarreal, and R. E. Mandrell. 2001. Wrinkled alfalfa seeds harbor more aerobic bacteria and are more difficult to sanitize than smooth seeds. J. Food Prot. 64:1292-1298. 10. Chen, H., D. G. Hoover, and D. H. Kingsley. 2005. Temperature and treatment time influence high hydrostatic pressure inactivation of feline calicivirus, a norovirus surrogate. J. Food Prot. 68:2389-2394.

139
11. Chen, H., D. Guan, and D. G. Hoover. 2006. Sensitivities of food-borne pathogens to pressure changes. J. Food Prot. 69:130-136. 12. Chen, H. 2007. Use of linear, Weibull, and log-logistic functions to model pressure inactivation of seven foodborne pathogens in milk. Food Microbiol. 24:197-204. 13. Chen, H. 2007. Temperature-assisted pressure inactivation of Listeria monocytogenes in turkey breast meat. Int. J. Food Microbiol. 117:55-60. 14. Fett, W. F., M. Tortorello, and T. Fu. 2005. Seed sprouts: the state of microbiological safety, p. 167-219. In K. R. Matthews (ed.), Microbiology of Fresh Produce. ASM Press, Washington, DC. 15. Ferguson, D. D., J. Scheftel, A. Cronquist, K. Smith, A. Woo-Ming, E. Anderson, J. Knutsen, A. K. De, and K. Gershman. 2005. Temporally distinct Escherichia coli O157 outbreaks associated with alfalfa sprouts linked to a common seed source - Colorado and Minnesota, 2003. Epidemiol. Infect. 133:439-447. 16. Gabriel, A. A. 2005. Microbial quality of chlorine soaked mung bean seeds and sprouts. Food Sci. Technol. Res. 11:95-100. 17. Gabriel, A. A., M. C. Berja, A. M. P. Estrada, M. G. A. A. Lopez, J. G. B. Nery, E. J. B. Villaflor. 2007. Microbiology of retail mung bean sprouts vended in public markets of National Capital Region, Philippines. Food Control. 18: 1307–1313. 18. Griggs, T. C. 2004. Alfalfa variety selection guidelines. Ag/Forage & Pasture/2004-02, Utah State University Cooperative Extension Fact Sheet. 19. Jaquette, C. B., L. R. Beuchat, and B. E. Mahon. 1996. Efficacy of chlorine and heat treatment in killing Salmonella Stanley inoculated onto alfalfa seeds and growth and survival of the pathogen during sprouting and storage. Appl. Environ. Microbiol. 62:2212-2215. 20. Kingsley, D. H., D. Guan, D. G. Hoover, and H. Chen. 2006. Inactivation of hepatitis A virus by high pressure processing: the role of temperature and pressure oscillation. J. Food Prot. 69:2454-2459. 21. Kural, A. G., and H. Chen. 2008. Conditions for a 5-log reduction of Vibrio vulnificus in oysters through high hydrostatic pressure treatment. Int. J. Food Microbiol. 122:180-187. 22. Kural, A. G., H. Chen, D. H. Kingsley, and A. E. H. Shearer. 2008. Conditions for high pressure inactivation of Vibrio parahaemolyticus in oysters. Int. J. Food Microbiol. 127:1-5. 23. Meyerowitz, S. 1999. My favorite greens. Better nutrition. 61:18-21.

140
24. MMWR. 1997. Outbreaks of Escherichia coli O157:H7 infection associated with eating alfalfa sprouts - Michigan and Virginia, June-July 1997. 46:741-744. 25. Mohle-Boetani, J. C., J. A. Farrar, S. B. Werner, D. Minassian, R. Bryant , S. Abbott , L. Slutsker , and D. J. Vugia. 2001. Escherichia coli O157 and Salmonella infections associated with sprouts in California, 1996-1998. Ann. Intern. Med. 135:239-247. 26. Munoz, M., B. D. Ancos, C. Sanchez-Moreno, and M. P. Cano. 2006. Evaluation of chemical and physical (high-pressure and temperature) treatments to improve the safety of minimally processed mung bean sprouts during refrigerated storage. J. Food Prot. 69:2395-2402. 27. Munoz, M., B. D. Ancos, C. Sanchez-Moreno, and M. P. Cano. 2007. Effects of high pressure and mild heat on endogenous microflora and on the inactivation and sublethal injury of Escherichia coli inoculated into fruit juices and vegetable soup. J. Food Prot. 70:1587-1593. 28. NACMCF. 1999. Microbiological safety evaluations and recommendations on sprouted 488 seeds. Int. J. Food Microbiol. 52:123-153. 29. Neetoo, H., M. Ye, and H. Chen. 2008. Potential application of high hydrostatic pressure to eliminate Escherichia coli O157:H7 on alfalfa sprouted seeds. Int. J. Food Microbiol. 128: 349-353. 30. Nienhuis, K. H., and H. J. Baltjes. 1985. Seed storage and germination in testing varieties for distinctness, uniformity and stability. Seed Sci. Technol. 13:19-25. 31. Pandrangi, S., M. W. Elwell, R. C. Anantheswaran, and L. F. LaBorde. 2003. Efficacy of sulfuric acid scarification and disinfectant treatments on eliminating Escherichia coli O157:H7 from alfalfa seeds prior to sprouting. J. Food Sci. 68:613-618. 32. Penas, E., C. Vidal-Valverde, J. Frias, and R. Gomez. 2008. Application of high-pressure treatment on alfalfa (Medicago sativa) and mung bean (Vigna radiata) seeds to enhance the microbiological safety of their sprouts. Food Control. 19:698-705. 33. Ponce, E., R. Pla, M. Capellas, B. Guamis, and M. Mor-Mur. 1998. Inactivation of Escherichia coli inoculated in liquid whole egg by high hydrostatic pressure. Food Microbiol. 15:265-272. 34. Ponce, E., R. Pla, E. Sendra, B. Guamis, and M. Mor-Mur. 1999. Destruction of Salmonella enteritidis inoculated in liquid whole egg by high hydrostatic pressure: comparative study in selective and non-selective media. Food Microbiol. 16:357-365.

141
35. Stewart, D., K. Reineke, J. Ulaszek, T. Fu, and M. Tortorello. 2001. Growth of Escherichia coli O157:H7 during sprouting of alfalfa seeds. Lett. Appl. Microbiol. 33:95-99. 36. Ting, E., V. M. Balasubramaniam, and E. Raghubeer. 2002. Determining thermal effects in high pressure processing. Food Technol. 56:31-35. 37. Weiss, A., and W. P. Hammes. 2005. Efficacy of heat treatment in the reduction of salmonellae and Escherichia coli O157:H7 on alfalfa, mung bean and radish seeds used for sprout production. Eur. Food Res. Technol. 221:187-191. 38. Winthrop, K. L. Alfalfa sprouts and Salmonella Kottbus infection: a multistate outbreak following inadequate seed disinfection with heat and chlorine. J. Food Prot. 66:13-17. 39. Wuytack, E. Y., C. W. Michiels, K. Meersseman, and A. M. Diels. 2003. Decontamination of seeds for seed sprout production by high hydrostatic pressure. J. Food Prot. 66:918-923.
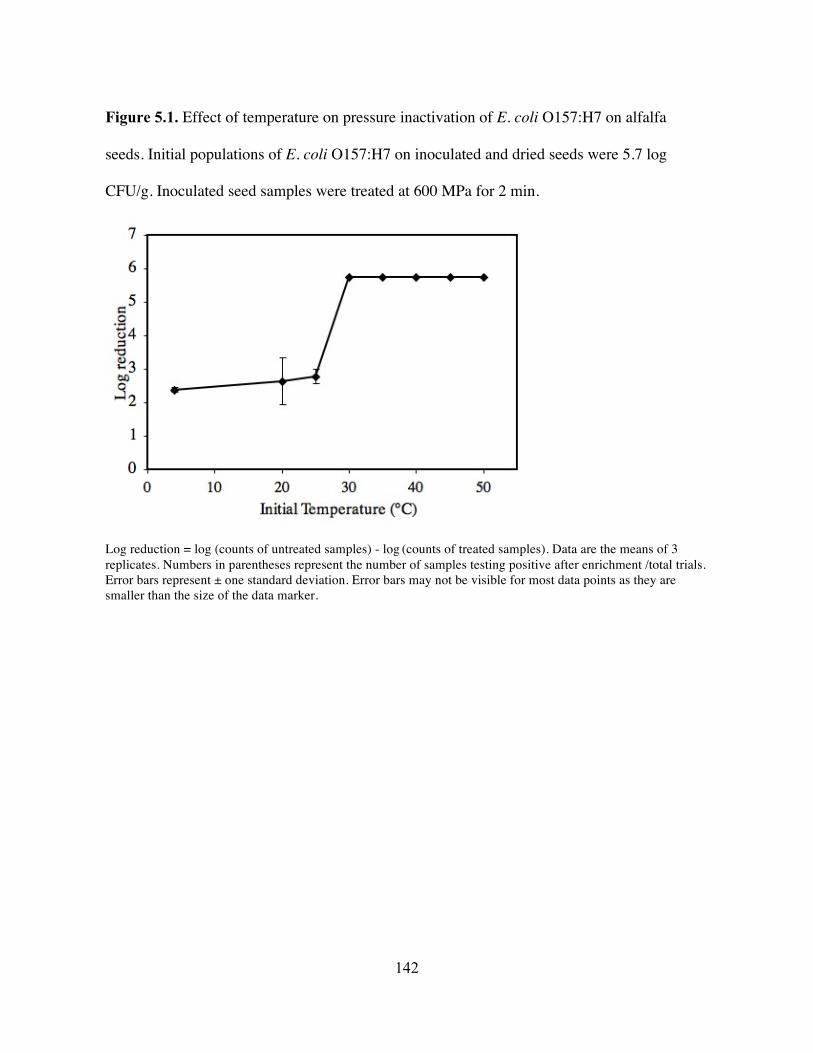
142
Figure 5.1. Effect of temperature on pressure inactivation of E. coli O157:H7 on alfalfa
seeds. Initial populations of E. coli O157:H7 on inoculated and dried seeds were 5.7 log
CFU/g. Inoculated seed samples were treated at 600 MPa for 2 min.
Log reduction = log (counts of untreated samples) - log (counts of treated samples). Data are the means of 3 replicates. Numbers in parentheses represent the number of samples testing positive after enrichment /total trials. Error bars represent ± one standard deviation. Error bars may not be visible for most data points as they are smaller than the size of the data marker.

143
Figure 5.2. Comparison of the germination rates of pressure-treated alfalfa seeds relative to
control (untreated) seeds. Error bars represent ± one standard deviation.

144
Figure 5.3. Comparison of the germination rates of fresh and aged alfalfa seeds pressure-
treated under various conditions relative to control (untreated) seeds. Error bars represent ±
one standard deviation. The y-axis scale was adjusted to a maximum of 110% to fit the error
bars into the chart area.

145
Figure 5.4. Effect of pressure-treatments carried out under various conditions on the average
sprout lengths of fresh and aged alfalfa seeds. Error bars represent ± one standard deviation.
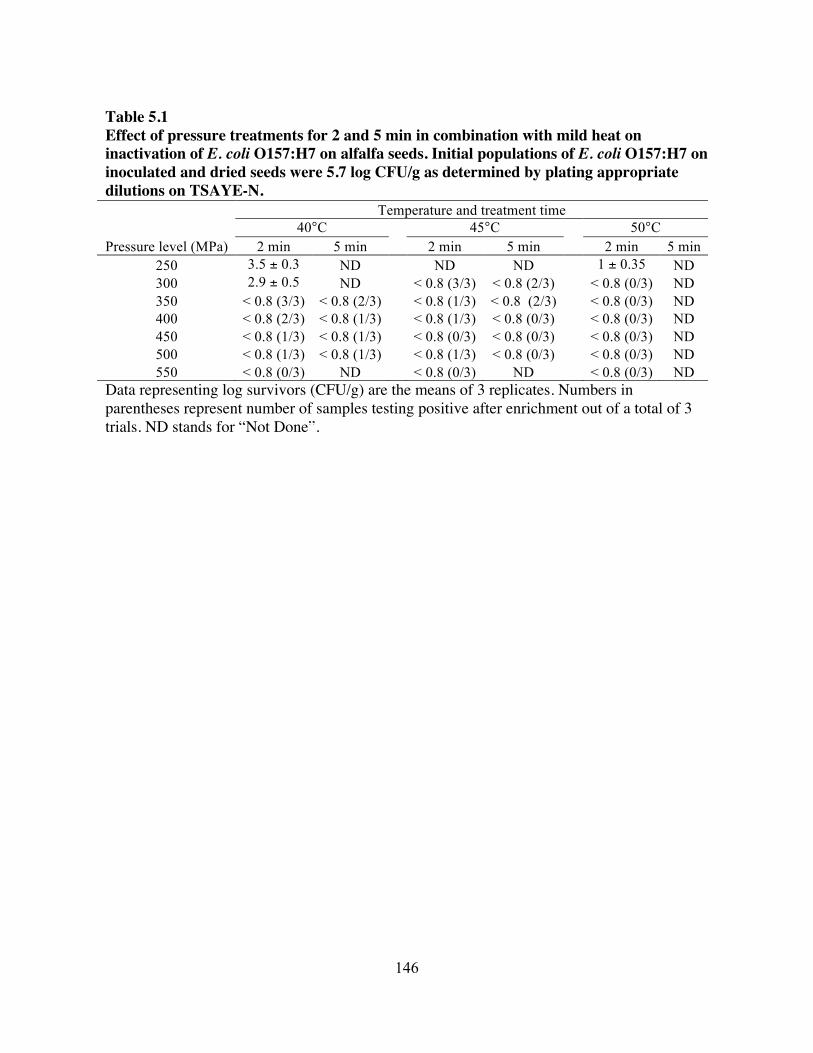
146
Table 5.1 Effect of pressure treatments for 2 and 5 min in combination with mild heat on inactivation of E. coli O157:H7 on alfalfa seeds. Initial populations of E. coli O157:H7 on inoculated and dried seeds were 5.7 log CFU/g as determined by plating appropriate dilutions on TSAYE-N.
Temperature and treatment time 40°C 45°C 50°C Pressure level (MPa) 2 min 5 min 2 min 5 min 2 min 5 min
250 3.5 ± 0.3 ND ND ND 1 ± 0.35 ND 300 2.9 ± 0.5 ND < 0.8 (3/3) < 0.8 (2/3) < 0.8 (0/3) ND 350 < 0.8 (3/3) < 0.8 (2/3) < 0.8 (1/3) < 0.8 (2/3) < 0.8 (0/3) ND 400 < 0.8 (2/3) < 0.8 (1/3) < 0.8 (1/3) < 0.8 (0/3) < 0.8 (0/3) ND 450 < 0.8 (1/3) < 0.8 (1/3) < 0.8 (0/3) < 0.8 (0/3) < 0.8 (0/3) ND 500 < 0.8 (1/3) < 0.8 (1/3) < 0.8 (1/3) < 0.8 (0/3) < 0.8 (0/3) ND 550 < 0.8 (0/3) ND < 0.8 (0/3) ND < 0.8 (0/3) ND
Data representing log survivors (CFU/g) are the means of 3 replicates. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials. ND stands for “Not Done”.

147
Chapter 6
Inactivation of Salmonella and Escherichia coli O157:H7 on artificially contaminated
alfalfa seeds using high hydrostatic pressure
Published in Food Microbiol. 2010. 27, 332-338.

148
Abstract
Alfalfa sprouts contaminated with Salmonella and Escherichia coli O157:H7 have
been implicated in several outbreaks of foodborne illnesses in recent years. The seed used for
sprouting appears to be the primary source of pathogens. Seed decontamination prior to
sprouting presents a unique challenge for the sprouting industry since cells of the pathogenic
survivors although undetectable after sanitizing treatments can potentially multiply back to
hazardous levels. The focus of this study was therefore to test the efficacy of high hydrostatic
pressure to eliminate a ~5 log CFU/g load of Salmonella and E. coli O157:H7 on alfalfa
seeds. Pressure treatment of 600 MPa for up to 25 min at 20°C could not result in complete
inactivation of Salmonella. High pressure treatment was then carried out either at sub-ambient
(4°C) or elevated (40, 45 and 50°C) temperatures to test the ability of high pressure to
eliminate Salmonella. Pressure treatment at 4 and 20°C did not deliver any satisfactory
inactivation of Salmonella while high pressure at elevated temperatures achieved complete
kill. Pre-soaking seeds prior to high pressure treatment also enhanced pressure inactivation of
Salmonella but at the expense of seed viability. High pressure treatment of 500 MPa for 2 min
at 45°C was able to eliminate wild-type Salmonella and E. coli O157:H7 strains without
bringing about any appreciable decrease in the seed viability.

149
Introduction
Sprouted seeds have a high nutritive value in addition to their anti-cholesterolemic and
anti-carcinogenic properties (Meyerowitz, 1999). In fact, consumption of raw seed sprouts has
become increasingly more popular among health conscious consumers. Unfortunately, this
has led to a concomitant increase in the incidence of sprout-associated foodborne illnesses
throughout the world (Fett and Cooke, 2003; Fett, 2006). Salmonella is the most frequently
reported cause of foodborne outbreaks of bacterial gastroenteritis in the United States after
Campylobacter and salmonellosis has been steadily increasing as a public health problem
since reporting began in 1943 (Tauxe, 1991). In North America alone, there have been at least
29 reports of sprout-borne Salmonella outbreaks, involving more than 1500 cases since 1995
(Taormina et al., 1999; Chapman, 2009). Salmonella appears to be linked more frequently to
alfalfa (Medicago sativa) sprout-related illnesses than other foodborne pathogens such as
Escherichia coli O157:H7 and Bacillus cereus (Proctor et al., 2001; Hora et al., 2007). Early
of 2009, a multi-state Salmonella outbreak linked to a variety of sprouts including alfalfa
sprouts sickened approximately 166 people. The cases were of Salmonella enterica serovar
Saintpaul strain while in 2008, Salmonella enterica serovar Typhimurium was involved in
another alfalfa sprouts outbreak (Chapman 2009; MMWR, 2009).
On the basis of epidemiological evidence as well as direct isolation, the seeds used for
sprouting have been found to be the most common source of these pathogens (Fett, 2006). As
a result, the U.S. Food and Drug Administration and the California Department of Health
Services currently recommends that sprout growers treat their seed with 20,000 ppm of free
chlorine for 15 min or with an equivalent approved antimicrobial treatment in addition to

150
testing spent irrigation water for the presence of Salmonella and E. coli O157:H7 (Waddell
and Troxell, 2000; Fett and Cooke, 2003). Although seed decontamination has improved the
safety of sprouted seeds, a number of sporadic outbreaks and product recalls incriminating
sprouts continue to occur. The limited performance of calcium hypochlorite along with the
fact that organic sprout growers are not allowed to treat seeds with such a high level of
chlorinated sanitizer has spurred a search for alternative seed decontamination treatments. In
addition, some treatments reduce sprouting efficacy, pose health hazards to workers or are
regarded by consumers as “unnatural” or “unhealthy” (Kumar et al., 2006). The challenge
encountered in disinfesting seeds is that pathogens can be located in deep cracks or crevices
on seed coats or in damaged areas thereby protecting them from antimicrobial treatments. If
pathogens are not completely eliminated, even low numbers of survivors can grow to
potentially hazardous levels on growing sprouts as a result of the favorable environmental and
nutritional conditions during the sprouting process (Holliday et al., 2001). Therefore, seed
decontamination strategies applied must ensure complete inactivation of human pathogens
while at the same time maintaining the viability and vigor of the seeds. This clearly indicates
the need for new strategies that are effective to control diverse Salmonella spp. on seeds since
it is known that multiple strains of Salmonella spp. have been implicated in sprout-related
disease outbreaks (NACMCF, 1999).
The application of HHP alone or in combination with mild heat have been studied
previously with reported varying degrees of efficacy (Ariefdjohan et al., 2004; Munoz et al.,
2006; Wuytack et al., 2003). Previous research in our laboratory has however shown that the
application of pressure at a level of 650 MPa for 15 min at 20°C (Neetoo et al., 2008) and 550

151
MPa for 2 min at 40°C (Neetoo et al., 2009) were adequate in completely inactivating E. coli
O157:H7 from alfalfa seeds whilst ensuring viability retention of the seeds. We also showed
that soaking seeds for an hour was critical in enhancing the pressure inactivation of E. coli
O157:H7 on seeds (Neetoo et al., 2009). Since the great majority of sprout-related illnesses
have been due to contamination with various serovars of Salmonella, the goal of the current
study was to therefore to develop a high-pressure process to simultaneously target a 5-log-
reduction of wild-type Salmonella and E. coli O157:H7 on alfalfa seeds to an undetectable
level (cells undetected by an enrichment method) without significantly affecting seeds
viability. The high pressure processing parameters were first optimized using nalidixic acid-
resistant Salmonella strains. The effect of the developed high pressure processes on the
viability of the seeds was then subsequently assessed. Finally, the ability of the processes to
eliminate wild-type Salmonella and E. coli O157:H7 strains was determined.
Materials and methods
Bacterial strains and preparation of inoculum.
Five Salmonella strains were used in this study: three S. Typhimurium strains, T43,
T45 and TDT 104, Salmonella enterica serovar Enteritidis E44 and Salmonella enterica
serovar Montevideo Mo57 (Courtesy of Dr. Joerger, University of Delaware). The cells of
Salmonella were adapted to grow in tryptic soy broth plus 0.6% yeast extract (Difco
Laboratories, Sparks, MD) supplemented with nalidixic acid to a final concentration of 50
µg/ml (Fisher Scientific, Hampton, NH) (TSBYE-N). Individual cultures were grown in
TSBYE-N overnight at 35°C. Cultures were then transferred (one loopful) into 10 ml of fresh
TSBYE-N and incubated at 35°C for 24 h. Equal volumes of individual cultures were mixed

152
to form a five-strain cocktail of Salmonella to a final cell density of 109 CFU/g.
Inoculation of seeds.
Appropriate dilutions of the five-strain culture cocktail (5 ml) was mixed with 75 ml
of sterile 0.1% peptone water (Fisher). Alfalfa seeds (Medicago sativa) (100 g), obtained from
International Specialty Supply (Cookeville, TN), were added to the cell suspension and gently
stirred for 5 min. The seeds were separated from the cell suspension by pouring the mixture
over a double layer of cheesecloth supported by a wire screen and dried inside a biosafety
hood at room temperature (21°C ± 1°C) for 24 h. Dried seeds with a water activity of 0.622
(AquaLab, Decagon Devices, Inc., WA)) and an approximate inoculation level of 105 CFU of
Salmonella per g were placed in sterile pouches and stored at 4°C.
High-pressure treatment.
Two grams of inoculated seeds and 3 ml of sterile deionized (DI) water were placed in
a 3-mil-thick pouch (nylon/polyethylene, Koch Supplies, Kansas City, MO). Seeds were
immersed in water during pressurization to enhance the baro-inactivation of pathogens and
viability retention of pressure-treated seeds as demonstrated previously by Neetoo el at.
(2008). To avoid leakage during pressure treatment, each sample pouch was placed in a larger
pouch of an 8-mil-thick polyvinyl chloride plastic (McMaster-Carr, Elmhurst, IL) and heat-
sealed. Pressure treatment of samples was immediately carried out using a high-pressure unit
with temperature control (model Avure PT-1; Avure Technologies, Kent, WA) and a
maximum operating pressure of 700 MPa. The experiments were conducted at 20°C (initial
seed sample temperature prior to pressure treatment) using water as a hydrostatic medium at a
pressure level of 600 MPa. The pressure increase rate was approximately 22 MPa/s. The

153
pressure release was almost immediate (< 4 s). Pressurization time reported in this study did
not include the pressure come up or release times.
Microbiological analysis.
Pouches containing treated seeds were cut open aseptically. The sample consisting of
the seed-water mixture was transferred into a stomacher bag to which 8 ml of sterile 0.1%
peptone water was added and subsequently stomached for 2 min at 260 rpm (Seward 400
Stomacher; Seward Medical Co., London, United Kingdom). The seed mixture was serially
diluted in sterile 0.1% peptone and surface-plated in duplicate on tryptic soy agar with 0.6%
yeast extract (Difco Laboratories, Sparks, MD) supplemented with nalidixic acid to a final
concentration of 50 µg/ml (TSAYE-N). TSAYE-N plates were incubated for 3 days at 35°C.
Presumptive colonies of Salmonella formed on the plates were enumerated. The seed mixture
was also directly enriched in 90 ml of TSBYE-N and incubated for 48 h at 35°C to allow
resuscitation of sublethally injured cells. Samples were streaked onto Xylose Lysine
Deoxycholate (XLD) agar (Difco Laboratories, Sparks, MD) plates supplemented with 50
mg/ml of nalidixic acid. After 24 h of incubation, the presence of black or black-centered
colonies typical of Salmonella was recorded as positive.
Application of low or elevated temperatures and/or extended treatment time on the
inactivation of Salmonella spp.
Two grams of inoculated seeds was mixed with 3 ml of sterile DI water, packaged,
and treated at 300 to 600 MPa for 2 min at 4, 20, 40, 45, and 50°C or at 300 to 600 MPa for 5
min at 40, 45 and 50°C as described previously. Samples were submerged in the water bath
surrounding the pressure cell for 10 min for samples to equilibrate to the water bath

154
temperature before pressurization. Immediately after pressure treatment, the samples were
cooled in an ice-water mixture. Samples were then microbiologically assayed as described
previously and enriched for the detection of survivors.
Effect of pre-soaking alfalfa seeds in water for varying times prior to pressure treatment
on inactivation of Salmonella spp.
Two g of inoculated seeds was soaked in 20 ml sterile DI-water for 0 (without
soaking), 15, 30 45 and 60 min at 20°C. At the end of the soaking period, the excess water
was subsequently decanted and seeds were placed into a pouch in the presence of 3 ml of
fresh sterile DI water and pressure-treated at 300-550 MPa for 2 min or 300-450 MPa for 5
min at 40°C as described above. Samples were then microbiologically assayed post-treatment
as described previously.
Germination test on selected treatments to determine optimal conditions for pressure
processing of seeds
To determine the effect of pressure treatment on the seed’s germination potential, two
grams of un-inoculated alfalfa seeds was mixed with 3 ml of DI water, packaged, and treated
at 600 MPa for 2 min at 40°C, 500 MPa for 2 min at 45°C, 500 MPa for 5 min at 40°C, 400
MPa for 5 min at 45°C and 350 MPa for 5 min at 50°C as described previously. In addition, 2
g of un-inoculated seeds were soaked for 45 min and pressure-treated at 500 MPa for 2 min at
40°C or soaked for 60 min, the excess water decanted and pressure-treated at 400 MPa for 5
min at 40°C as described previously. Untreated (control) and pressure-treated seeds were then
soaked in DI water after pressure-treatment for an overall soaking time of 3 h prior to setting
them to germinate. One hundred seeds were drawn from the soaked seeds and spread evenly

155
on pieces of wet paper towels on a plastic rack, which in turn was placed in a water-filled
bucket to provide a moist environment for the seeds. The water level was maintained below
the level of seeds. The bucket was kept at room temperature for 8 days (suggested by the seed
provider) and misted daily. The bucket was covered loosely with a piece of plastic film to
allow exchange of air between the inside and outside of the bucket. Sprouted seeds
characterized by the emergence of the root tip (radicle) were enumerated 3 to 8 days after the
germination system was set up, and the values were recorded as percent germination. The
incubation duration was extended to up to 8 days to give sufficient time for seeds with a
delayed sprouting onset to germinate.
Assessment of the robustness of selected treatments with respect to complete elimination
of wild-type Salmonella and E. coli O157:H7
Two grams of alfalfa seeds inoculated with wild-type Salmonella was mixed with 3 ml
of DI water, packaged, and treated at 600 MPa for 2 min at 40°C, 500 MPa for 2 min at 45°C
or 500 MPa for 5 min at 40°C. Samples were then pre-enriched in Lactose broth (90 ml) for
24 h at 35°C. After the pre-enrichment step, 0.1 ml of the mixture was transferred to 10 ml
Rappaport-Vassiliadis (RV) medium and incubated for another 24 h. After the secondary
enrichment step, the sample was then streaked onto XLD plates (U.S. FDA, 2007). Wild-type
E. coli O157:H7 strains, Strains 250, 251 and 1730, Cider strain and J58, were also used
(Neetoo et al., 2008). Two grams of alfalfa seeds inoculated with a cocktail of the five wild-
type E. coli O157:H7 strains were mixed with 3 ml of DI water, packaged, and treated at 500
MPa for 2 min at 45°C or 500 MPa for 5 min at 40°C as described previously. Samples were
then enriched in 90 ml of EHEC Enrichment Broth (EEB) for 24 h at 35°C. After the

156
enrichment step, the sample was then streaked onto CT-SMAC plates and incubated for 24 h
(U.S. FDA, 2002).
Testing of sprouted seed samples and waste irrigation water for surviving
pathogens
Two grams of seeds inoculated with a ~ 5 log burden of wild-type Salmonella or E.
coli O157:H7 were pressure treated at 500 MPa at 45°C for 2 min as described previously.
Treated seeds were then soaked for 3 h at ambient temperature in sterile DI water. After
soaking, the water was then decanted and the seeds were placed onto a pre-wet Whatman
filter paper in sterile plastic boxes and incubated at ambient temperature for 2 days. During
the time of germination, sterile DI water was sprayed onto the sprouting seeds daily. After 48
h, sprouted and non-sprouted seeds were scooped and transferred into sterile stomacher bags,
enriched and analyzed as described above. In addition, 10 ml of the spent water was collected
at the bottom of the plastic container and similarly tested. The presence of growth on CT-
SMAC and XLD plates were further confirmed for identification using the RapidChek® tests
for E. coli O157 and Salmonella respectively (Strategic Diagnostics Inc., Newark, DE, USA).
Statistical analysis.
All experiments were replicated three times. Where appropriate, statistical analyses
were conducted using Minitab release 15 (Minitab Inc., University Park, PA). One-way
analysis of variance and Tukey’s one-way multiple comparisons were used to determine
differences in the populations of Salmonella O157:H7 recovered on treated alfalfa seeds and
differences in the germination rates of seeds. Differences were considered statistically
significant at the 95% confidence level (P < 0.05).

157
Results and discussion
Determination of the pressure inactivation curve of Salmonella spp. on alfalfa seeds
Survival curve of a five-strain Salmonella cocktail on seeds pressurized at 20°C is
shown in Fig. 6.1. Pressure-treatment for 5-25 min significantly (P < 0.05) reduced the
population and an increase in treatment time was accompanied by a progressively higher
degree of inactivation, achieving a maximum reduction of 4.5 log CFU/g of Salmonella. The
inactivation curve exhibited a nearly linear decline in surviving cells with time, marked by a
steady drop in bacterial counts, in contrast to the more frequently observed pressure
inactivation curve characterized by a rapid initial drop followed by tailing caused by a
diminishing inactivation rate. Such a linear survivor curve suggests that the population was
rather homogeneous in terms of its pressure resistance.
Pressure inactivation of Salmonella has been widely studied at ambient temperature.
Patterson et al. (2005) showed that a pressure treatment of 350 MPa and 15 min at 20°C
reduced S. Typhimurium by 5 log cycles. Chen et al. (2006) demonstrated a 6.5-log reduction
of S. Enteritidis at 600 MPa for 10 min at 21.5°C in UHT whole milk. Tholozan et al. (2000)
exposed S. Typhimurium Mutton (ATCC 13311) in sodium citrate (pH 5.6) and sodium
phosphate (pH 7.0) buffers to 400 MPa for 10 min and 20°C for complete inactivation (> 8
log reduction). Guan et al. (2005) showed that pressure treatment of 450 MPa for at least 30
min or 20 min at 500 and 550 MPa at ambient temperature brought about a 5.0-log reduction
of S. Typhimurium DT 104 in UHT milk. In general, inactivation curves did not follow first-
order kinetics but rather tended to be exponential, with an initial rapid decrease in numbers
followed by a “tail” indicating that a small fraction of the population was highly baro-resistant

158
(Chen, 2007). In addition, as several authors already mentioned, the composition and water
activity of the substrate in which the microorganisms are suspended are also likely factors that
affect their high-pressure tolerance (Oxen and Knorr, 1993; Palou et al., 1997; Patterson and
Linton, 2008).
Patterson (2008) noted that significant variation in pressure resistance has been
reported among different strains of the same genus which may be attributed to differences in
susceptibility to membrane damage, although the exact nature of the lethal effect and the role
of the membrane structure still have to be clarified. Chen et al. (2009) observed that when
thirty strains of Listeria monocytogenes were exposed to a treatment of 400 MPa for 2 min at
21°C, the strains exhibited reductions ranging from 1.9 to 7.1 log CFU/ml. In recognition of
the natural variability and uniqueness of strains, it is preferable to use a cocktail of strains in
inactivation and validation studies conducted on food (Balasupramaniam et al., 2004). Hence,
in our challenge studies, a composite of five strains of Salmonella were used to that effect. S.
Typhimurium DT 104 was included as part of the five-strain cocktail since this strain has been
the object of rising alarm, spreading at a fast pace through Britain and the United States over
the past 10 to 15 years (Keene, 1999).
In our previous studies, we demonstrated that pressure treatment of 600 MPa on
alfalfa seeds inoculated with a 5 log population of a cocktail of five nalidixic acid-resistant E.
coli O157:H7 strains resulted in 3-5 log reductions when the pressure exposure time was from
2-20 min; however, complete elimination was not possible at 600 MPa (Neetoo et al., 2008).
In the current study, when pressure treatments at 600 MPa were applied on a five-strain
Salmonella cocktail, the latter displayed a higher degree of pressure resistance with log

159
reductions ≤ 4.5 log CFU/g after 25 min exposure to pressure. Recently, Barak et al. (2002)
demonstrated the survival advantage of S. enterica over E. coli O157:H7 for attachment to
sprouting seeds, sprouts and plant tissue in general. The authors explained that the difference
in attachment could particularly be explained by the distinct ability of S. enterica to produce
aggregative fimbriae (curli). It is thought that S. enterica have a higher tendency to express
curli that play an essential role in the attachment of the pathogen to seeds and plant tissue in
general. It is possible that the higher pressure resistance of Salmonella may be attributed to
the stronger attachment to the seed tissue hence affording a certain degree of protection
against pressure inactivation.
Application of low or elevated temperatures and/or extended treatment time on the
inactivation of Salmonella
Although high pressure treatment represents a more costly process than conventional
seed decontamination methods, we wanted to investigate whether the processing parameters
could be manipulated to influence the outcome and economics of the high-pressure treatment.
To achieve more than 5 log cycles reduction of Salmonella on seeds, higher pressure (> 600
MPa) and/or extended treatment time (> 25 min) could be used. However, the upper pressure
limit for commercial high pressure applications is 600 MPa and long pressurization time is
not economically feasible. The effect of treatment temperature is therefore of great practical
interest, because combined pressure-temperature processing may deliver greater microbial
inactivation ratios while operating at lower pressure levels and/or for shorter periods of time.
The economical advantage of combining pressure with temperature would also result in a less
costly equipment at a higher throughput, with a considerable decrease in the production cost

160
(per mass unit of food) (Van den Berg et al., 2001).
Hence, our next step was to investigate the ability of high pressure to completely
eliminate Salmonella spp. on alfalfa seeds subjected to various pressure/time/temperature
combinations. Results for the pressure inactivation of Salmonella on alfalfa seeds at different
initial sample temperatures are shown in Table 6.1. The enhancement of temperature on
pressure inactivation of Salmonella is clearly evident marked by the reduction to below
detectable limits of the initial inoculum load under various ternary combinations of
pressure/time/temperature. The most effective treatment was observed at 50°C, followed by
45 and 40°C. The degree of inactivation increased with increasing pressure with lower
survival rates at 600 MPa than at 300 MPa for all temperatures tested.
When the initial samples temperatures were set to 4 or 20°C, Salmonella spp.
responded quite similarly to pressures ranging from 300-600 MPa with no statistically
significant enhancement (P < 0.05) at a sub-ambient temperature, achieving a maximum
reduction of 1.5 and 2.0 log CFU/g at 600 MPa at 4 and 20°C, respectively. Previous studies
have shown that the combination of pressure and low or subzero treatment temperatures can
be more effective at killing microorganisms than treatment at ambient temperature (20°C).
Lopez-Caballero et al. (2002) found greater microbial inactivation when the pressure
treatment was carried out at refrigeration temperatures rather than at ambient temperature.
They reported 1- to 2-log greater inactivation of Pseudomonas fluorescens at 5°C than with
pressure treatment at 20 or 35°C. When L. innocua was pressure-treated at 300 MPa for 15
min in liquid whole egg, a 1.5 to 2 log inactivation was achieved at -15 or 2°C, compared
with virtually no inactivation at 20°C (Ponce et al., 1998). Kural and Chen (2008) also found

161
that cold temperature enhanced pressure inactivation of Vibrio vulnificus when compared with
treatment at ambient temperature. However, we did not observe any enhancement in pressure
inactivation of Salmonella at low temperatures compared to ambient temperatures,
demonstrating that the effect of sub-ambient temperature on pressure inactivation of
microorganisms are strain-dependent.
When the initial sample temperature was 40°C and higher, inactivation was
significantly higher (P < 0.05) and more pressure-dependent, achieving > 5 log reductions at ≥
300 MPa (50°C), ≥ 400 MPa (45°C) and ≥ 450 MPa (40°C) although survivors were still
detected post-enrichment. However, survivors were not detected using the enrichment method
when the pressure was increased to 600 MPa (40°C) and 500 MPa (≥ 45°C). Increasing the
treatment temperature from 40 through 50°C greatly enhanced the pressure inactivation of
Salmonella reducing the pressure required to completely eliminate the pathogen. Bacterial
cells are known to be relatively less barosensitive at 20-35°C, but become more sensitive to
pressurization above 35°C, due to phase transition of membrane lipids (Kalchanyanad et al.,
1998a, b; Ludwig et al., 2002).
Since the findings showed that a 2-min application of high pressure at a level of 300-
450 MPa at 40 and 45°C resulted in > 5 log inactivation, the pressure treatment time was thus
extended to 5 min to investigate whether a lower pressure level could result in an equivalent
kill in an attempt to reduce the treatment pressure level. When the pressure exposure time was
extended to 5 min, as anticipated, a lower pressure bracket (300-500 MPa) brought about an
equivalent kill. Specifically, exposure to 500 MPa at 40°C or ≥ 400 MPa at 45°C or ≥ 350
MPa at 50°C for 5 min completely eliminated a 5 log load of Salmonella.

162
Effect of pre-soaking alfalfa seeds in water for varying times prior to pressure treatment
on inactivation of Salmonella
To further reduce the pressure level needed for complete elimination of Salmonella on
seeds, the effect of pre-soaking of seeds on pressure inactivation of Salmonella was
determined. Tables 3 and 4 show that treatments at 300-600 MPa without prior soaking
brought about 2.8-5.8 log CFU/g reduction at 40°C. When seeds were soaked for > 30 min, a
significant enhancement (P < 0.05) in pressure inactivation of Salmonella relative to un-
soaked seeds was observed. In addition, a direct relationship between the degree of
inactivation and the soaking time as evidenced by larger population reductions was observed
across the entire pressure spectrum (300-550 MPa) investigated. Seeds soaked for > 45 min
followed by pressure-treatment at > 500 MPa at 40°C for 2 min produced no detectable
counts and tested negative after enrichment (Table 6.3). When the pressure holding time was
extended to 5 min, a pressure level of > 400 MPa was adequate to completely decontaminate
seeds with a minimum soaking time of 60 min (Table 6.4). Again, the same trend was
observed i.e the longer the soaking time, the greater the degree of pressure inactivation.
Previously, Delaquis et al. (1999) performed a microscopic examination of alfalfa
seeds and reported that although the seed surface is relatively smooth, the stem scar is
relatively porous with areas capable of harboring pathogens, thus affording protection to
hidden bacterial cells. In addition to its topographic complexity, the surface of alfalfa seeds is
covered with a waxy cuticle (cutin) thus lowering their water wettability. Charkowski et al.
(2001) however mentioned that by pre-soaking of seeds in water for a certain period of time,
water is believed to permeate the seed coat causing bacteria trapped in cracks, crevices or

163
other discontinuities to be released. Indeed, we demonstrated that when seeds were pressure-
treated whilst immersed in water with no prior soaking, water did not really have time to
permeate the seed coat within the short pressurization time frame (2-5 min) investigated in
our study. By contrast, seeds pre-soaked in water for a long enough time allowed an un-
inhibited penetration of water into the seeds such that an hour of imbibition was adequate to
allow water to access the deepest cracks or crevices of the seed coat (Neetoo et al., 2009b).
This observation is made in the current study and is also congruent with previous findings
reported by the same author on the pressure-inactivation of E. coli O157:H7 on un-soaked and
soaked alfalfa seeds (Neetoo et al., 2009). It is also thought that the pre-soaking step may be
important in raising the water activity of the seeds which is known to have a bearing on the
pressure inactivation of bacterial cells. It is well documented that the higher the water activity,
the more enhanced is the pressure inactivation of microorganisms (Oxen and Knorr, 1993;
Palou et al., 1997; Goodridge et al., 2006; Kingsley and Chen, 2008). The critical role played
by water in pressure inactivation of E. coli O157:H7 on seeds was also demonstrated by
Neetoo et al. (2008; 2009). Hence it can be concluded that there is an intimate interplay
between the soaking time, pressure magnitude and treatment temperature which are critical
factors in ensuring the efficacy of HHP to decontaminate alfalfa seeds.
Germination test on selected treatments to determine optimal conditions for pressure
processing of seeds
Results from section 3.2 and 3.3 have shown that complete inactivation of Salmonella
spp. is possible when seeds are pressure-treated under selected conditions of pressure
exposure time, temperature, pressure level and soaking time. The germination percentages as

164
determined 3-8 days from the onset of germination showed that seeds pressure-treated under
the different conditions were affected to variable extents depending on the conditions of the
treatment (Table 6.5.). The treatments at 600 MPa at 40°C for 2 min, 500 MPa at 40°C for 5
min, 500 MPa at 45°C for 2 min and 400 MPa at 45°C for 5 min were the most promising as
they had no significant adverse effects (P < 0.05) on the seeds’ viability achieving a
germination percentage of ≥ 95% after 8 days of germination. Pressure-treatments on un-
soaked alfalfa seeds at 40 and 45°C brought about greater viability retention than when the
temperature was set to 50°C.
As far as soaked seeds are concerned, it is not clear what mechanisms are responsible
for the lower germination yields but it is possible that denaturation of proteinaceous
molecules such as enzymes might have occurred during pressurization following their
activation during prolonged water imbibition. It is also likely that during soaking, seeds
uptake water and as a result of the increased moisture content, the seeds advance into an
active state whereby physiological changes associated with the initial stages of germination
start to take place (Neetoo et al., 2009).
Assessment of the robustness of selected treatments with respect to complete elimination
of wild-type Salmonella and E. coli O157:H7
The three optimum treatments determined in Section 3.4 were able to completely
inactivate Salmonella nalidixic-acid resistant mutants; however only two treatments, 500 MPa
for 5 min at 40°C and 500 MPa for 2 min at 45°C were able to consistently eliminate wild-
type Salmonella strains on alfalfa seeds. Further testing on wild-type E. coli O157:H7 strains
showed that pressure-treatment at 500 MPa for 2 min at 45°C was the only treatment that

165
could achieve a complete inactivation. Since seeds pressure-treated under 500 MPa at 45°C
for 2 min achieved > 95% germination rate (Table 6.5.), this study thus indicates that high
pressure applied in conjunction with mild heat can be an effective method for inactivating
Salmonella and E. coli O157:H7 on alfalfa seeds, hence pointing to the plausibility of
pressure application for effective control of enteric pathogens on alfalfa seeds.
Testing of sprouted seed samples and waste irrigation water for surviving pathogens
Overall, sprouted seeds and spent irrigation water samples inoculated with E. coli
O157:H7 and pressure treated at 500 MPa and 45°C for 2 min produced bright pink growth on
CT-SMAC in out of three cases. Seeds and collected water samples initially inoculated with
Salmonella produced bright yellow growth on XLD plates in two out of three trials. Since
colony morphology and color were atypical of E. coli O157:H7 and Salmonella respectively,
we confirmed the absence of both pathogens in all cases using immunoassay-based tests.
Conclusions
In conclusion, this study demonstrated that the pressure inactivation of Salmonella
including S. Typhimurium DT 104 on alfalfa seeds followed a fairly linear rate of
inactivation. Results clearly indicate that destruction of Salmonella spp. on alfalfa seeds
depended on pressure and the temperature at which it was applied. Also, inactivation was less
pronounced at low and room temperatures while higher temperatures gave better results. High
pressure (500 MPa) combined with mild heating (45°C) could be an effective means to
decontaminate seeds whilst retaining seed viability. The inactivation data garnered in this
study should be invaluable for seed processing for safety, especially as exemplified using S.
typhimurium DT 104. With the ever expanding applications of HHP for the preservation and

166
processing of food, reliable determination of optimum processing parameters, with regard to
temperature, time and pressure magnitude, will become critical process factors towards
ensuring the safety of pressure-treated foods.
Acknowledgement
This study was supported by a start-up fund from the Department of Animal and Food
Sciences at the University of Delaware.

167
References Balasubramaniam, V.M., Ting, E.Y., Stewart, C.M., Robbins, J.A., 2004. Recommended laboratory practices for conducting high-pressure microbial experiments. Innov. Food Sci. Emerg. 5, 299-306. Barak, J.D., Whitehand, L.C., Charkowski, A.O., 2002. Differences in attachment of Salmonella enterica serovars and Escherichia coli O157:H7 to alfalfa sprouts. Appl. Environ. Microbiol. 68, 4758-4763. Charkowski, A.O., Sarreal, C.Z., Mandrell, R.E., 2001. Wrinkled alfalfa seeds harbor more aerobic bacteria and are more difficult to sanitize than smooth seeds. J. Food Prot. 64, 1292-1298. Chapman, B., 2009. Sprouts associated outbreaks in North America, 1990-2009. Ben Chapman Bites, Food Safety Network. Viewed 05/14/ 2009 from http://foodsafety.k-state.edu/en/article-details.php?a=2&c=6&sc=36&id=865 Chen, H., Guan, D., Hoover, D.G. 2006. Sensitivities of foodborne pathogens to pressure changes. J. Food Prot. 69, 130-136. Chen, H., 2007. Use of linear, Weibull, and log-logistic functions to model pressure inactivation of seven foodborne pathogens in milk. Food Microbiol. 24, 197-204. Chen, H., Neetoo, H., Ye, M., Joerger, R.D., 2009. Differences in pressure tolerance of Listeria monocytogenes strains are not correlated with other stress tolerances and are not based on differences in CtsR. Food Microbiol., 26, 404-408. Delaquis, P.J., Sholberg, P.L., Stanich, K., 1999. Disinfection of mung bean seed with gaseous acetic acid. J. Food Prot. 62, 953-957. Fett, W.F., Cooke, P.H., 2003. Reduction of Escherichia coli O157:H7 and Salmonella on laboratory-inoculated alfalfa seed with commercial citrus-related products. J. Food Prot. 66, 1158-1165. Fett, W.F., 2006. Interventions to ensure the microbial safety of sprouts. In: Sapers, G.M., Gorny, J.R., Yousef, A.E. (Eds.), Microbiology of Fruits and Vegetables. Taylor and Francis Group, LLC, Boca Raton, FL., pp. 187-210. Goodridge, L.D., Willford, J., Kalchayanand, N., 2006. Destruction of Salmonella Enteriditis inoculated onto raw almonds by high hydrostatic pressure. Food Res. Int. 39, 408-412.

168
Guan, D., Chen, H., Hoover, D.G., 2005. Inactivation of Salmonella Typhimurium DT 104 in UHT whole milk by high hydrostatic pressure. Int. J. Food Microbiol. 104, 145-153. Holliday, S.L., Scouten, A.J., Beuchat, L.R., 2001. Efficacy of chemical treatments in eliminating Salmonella and Escherichia coli O157:H7 on scarified and polished alfalfa seeds. J. Food Prot. 64, 1489-1495. Hora, R., Kumar, M., Kostrzynska, M., Dixon, M.A., Warriner, K., 2007. Inactivation of Escherichia coli O157:H7 and Salmonella on artificially or naturally contaminated mung beans (Vigna radiata L) using a stabilized oxychloro-based sanitizer. Lett. Appl. Microbiol. 44, 188-193. Kalchayanand, N., Sikes, A., Dunne, C.P., Ray, B., 1998. Factors influencing death and injury of foodborne pathogens by hydrostatic pressure-pasteurization. Food Microbiol. 15, 207-214. Kalchayanand, N., Sikes, A., Dunne, C.P., Ray, B., 1998. Interaction of hydrostatic pressure, time and temperature of pressurization and pediocin AcH on inactivation of foodborne bacteria. J. Food Prot. 61, 425-431. Keene, W.E., 1999. Lessons from investigations of foodborne disease outbreaks. J. Am. Med. Assoc. 281, 1845-1847. Kingsley, D.H., Chen, H., 2008. Aqueous matrix compositions and pH influence feline calicivirus inactivation by high pressure processing. J. Food Prot. 71, 1598-1603. Kumar, M., Hora, R., Kostrzynska, M., Waites, W.M., Warriner, K., 2006. Inactivation of Escherichia coli O157:H7 and Salmonella on mung beans, alfalfa, and other seed types destined for sprout production by using an oxychloro-based sanitizer. J. Food Prot. 69, 1571-1578. Kural, A., Chen, H., 2008. Conditions for a 5-log reduction of Vibrio vulnificus in oysters through high hydrostatic pressure treatment. Int. J. Food Micro. 122, 180-187. Lopez-Caballero, M.E., Carballo, J., Jimenez-Colmenero, F., 2002. Microbial inactivation in meat products by pressure/temperature processing. J. Food Sci. 67, 797-801. Ludwig, H., 2002. Cell biology and high pressure: applications and risks. Int. J. Biochem. Bio. 1595, 390-391. Meyerowitz, S., 1999. My favorite greens. Better nutrition. 61, 18-21.
MMWR, 2009. Outbreak of Salmonella serotype Saintpaul infections associated with eating alfalfa sprouts – United States, 2009. Morb. Mortal. Wkly. Rep. 58, 1-4.

169
NACMCF (National Advisory Committee on Microbiological Criteria for Foods), 1999. Microbiological safety evaluations and recommendations on sprouted seeds. Int. J. Food Microbiol. 52, 123-153. Neetoo, H., Chen, H., Ye, M., 2008. Potential application of high hydrostatic pressure to eliminate Escherichia coli O157:H7 on alfalfa sprouted seeds. Int. J. Food Microbiol. 128, 348-353. Neetoo, H., Chen, H., Pizzolato, T., 2009. Elimination of Escherichia coli O157:H7 from alfalfa seeds through a combination of high hydrostatic Pressure and Mild Heat. Appl. Env. Microbiol. 75, 1901-1907. Neetoo, H., Ye, M., Chen, H., 2009. Factors affecting the efficacy of pressure inactivation of Escherichia coli O157:H7 on alfalfa seeds and seed viability. Int. J. Food Microbiol. 131, 218-23. Oxen, P., Knorr, D., 1993. Baroprotective effects of high solute concentrations against inactivation of Rhodotorula rubra. Lebensm. Wiss. Technol. 26, 220-223. Palou, E., Lopez-Malo, A., Barbosa-Canovas, G. V., Welti-Chanes, J., Swanson, B. G., 1997. Effect of water activity on high hydrostatic pressure inhibition of Zygosaccharomyces bailii. Lett. Appl. Microbiol. 24, 417-420. Patterson, M.F., 2005. Microbiology of pressure-treated foods. J. Appl. Microbiol. 98, 1400-1409. Patterson, M.F., Linton, M., 2008. Factors affecting inactivation of foodborne bacteria by high pressure. In: Michiels, C., Bartlett, D.H., Aertsen, A., (Eds.), High Pressure Microbiology, ASM Press, Washington, DC, pp. 181–195. Ponce, E., Pla, R., Mor-Mur, M., Gervilla, R., Guamis, B., 1998. Inactivation of Listeria innocua inoculated in liquid whole egg by high hydrostatic pressure. J. Food Prot. 61, 119-122. Proctor, M.E., Hamacher, M., Tortorello, M.L., Archer, J.R., Davis, J.P., 2001. Multistate outbreak of Salmonella serovar Muenchen infections associated with alfalfa sprouts grown from seeds pretreated with calcium hypochlorite. J. Clin. Microbiol. 39, 3461-3465. Taormina, P.J., Beuchat, L.R., 1999. Behavior of enterohemorrhagic Escherichia coli O157:H7 on alfalfa sprouts during the sprouting process as influenced by treatments with various chemicals. J. Food Prot. 62, 850-856. Tauxe, R.V., 1991. Salmonella: A postmodern pathogen. J. Food Prot. 54, 563-568.

170
Tholozan, J.L., Ritz, M., Jugiau, F., Federighi, M., Tissier, J.P., 2000. Physiological effects of high hydrostatic pressure treatments on Listeria monocytogenes and Salmonella Typhimurium. J. Appl. Microbiol. 88, 202-212. U.S. Food and Drug Administration. Bacteriological Analytical Manual, 2002, Ch. 4a. http://www.fda.gov/Food/ScienceResearch/LaboratoryMethods/BacteriologicalAnalyticalManualBAM/ucm070080.htm (Last accessed 07/29/09). U.S. Food and Drug Administration. Bacteriological Analytical Manual, 2007, Ch. 5. http://www.fda.gov/Food/ScienceResearch/LaboratoryMethods/BacteriologicalAnalyticalManualBAM/ucm070149.htm (Last accessed 07/29/09). Van den Berg, R.W., Hoogland, H., Lelieveld, H.L.M., Van Schepdael, L., 2001. High pressure equipment designs for food processing applications. In: Hendricks, M.E.G., Knorr, D. (Eds.), Ultra High Pressure Treatments of Foods. Kluwer Academic/Plenum Publishers, New York, NY, pp. 297-312. Waddell, J.M, Troxell, T.C., 2000. Safer processing of sprouts. U.S. FDA and the California Department of Health Services, Food and Drug Branch, available at http://vm.cfsan.fda.gov/˜dms/sprouvid.html

171
Figure 6.1. Pressure inactivation curve of nalidixic acid-resistant strains of Salmonella spp.
on alfalfa seeds treated at 600 MPa and 20°C. Error bars represent ± one standard deviation.
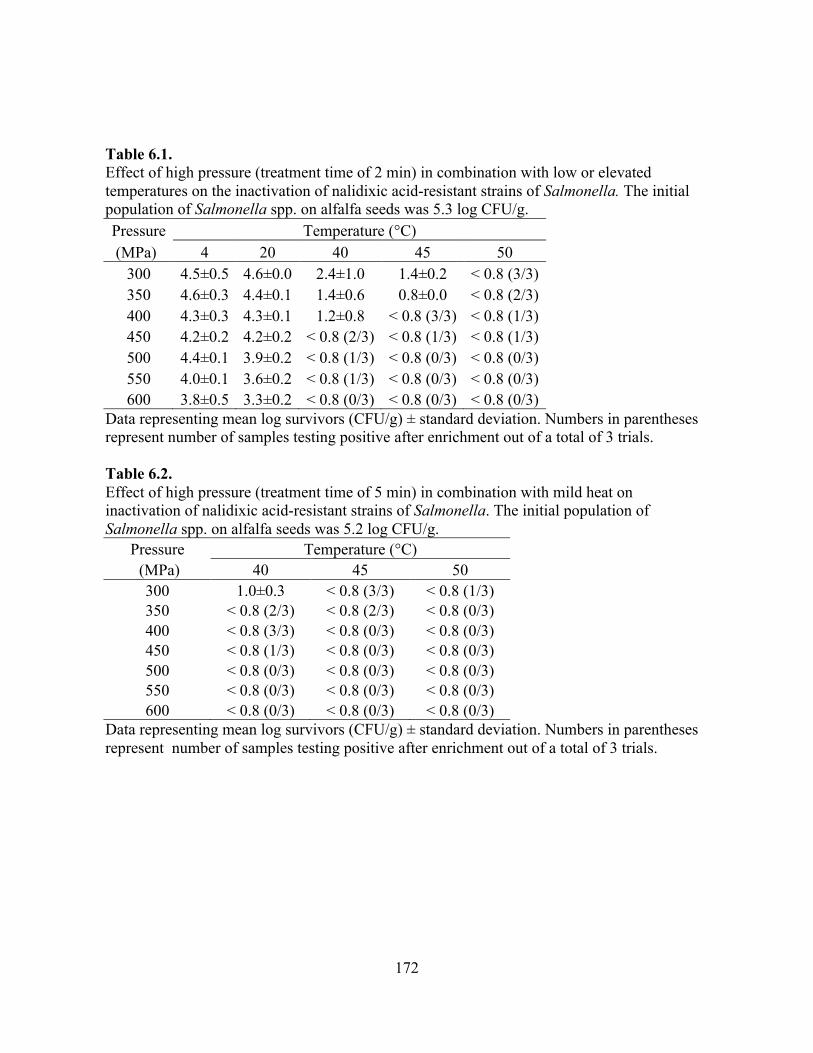
172
Table 6.1. Effect of high pressure (treatment time of 2 min) in combination with low or elevated temperatures on the inactivation of nalidixic acid-resistant strains of Salmonella. The initial population of Salmonella spp. on alfalfa seeds was 5.3 log CFU/g. Pressure Temperature (°C) (MPa) 4 20 40 45 50
300 4.5±0.5 4.6±0.0 2.4±1.0 1.4±0.2 < 0.8 (3/3) 350 4.6±0.3 4.4±0.1 1.4±0.6 0.8±0.0 < 0.8 (2/3) 400 4.3±0.3 4.3±0.1 1.2±0.8 < 0.8 (3/3) < 0.8 (1/3) 450 4.2±0.2 4.2±0.2 < 0.8 (2/3) < 0.8 (1/3) < 0.8 (1/3) 500 4.4±0.1 3.9±0.2 < 0.8 (1/3) < 0.8 (0/3) < 0.8 (0/3) 550 4.0±0.1 3.6±0.2 < 0.8 (1/3) < 0.8 (0/3) < 0.8 (0/3) 600 3.8±0.5 3.3±0.2 < 0.8 (0/3) < 0.8 (0/3) < 0.8 (0/3)
Data representing mean log survivors (CFU/g) ± standard deviation. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials. Table 6.2. Effect of high pressure (treatment time of 5 min) in combination with mild heat on inactivation of nalidixic acid-resistant strains of Salmonella. The initial population of Salmonella spp. on alfalfa seeds was 5.2 log CFU/g.
Pressure Temperature (°C) (MPa) 40 45 50
300 1.0±0.3 < 0.8 (3/3) < 0.8 (1/3) 350 < 0.8 (2/3) < 0.8 (2/3) < 0.8 (0/3) 400 < 0.8 (3/3) < 0.8 (0/3) < 0.8 (0/3) 450 < 0.8 (1/3) < 0.8 (0/3) < 0.8 (0/3) 500 < 0.8 (0/3) < 0.8 (0/3) < 0.8 (0/3) 550 < 0.8 (0/3) < 0.8 (0/3) < 0.8 (0/3) 600 < 0.8 (0/3) < 0.8 (0/3) < 0.8 (0/3)
Data representing mean log survivors (CFU/g) ± standard deviation. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials.
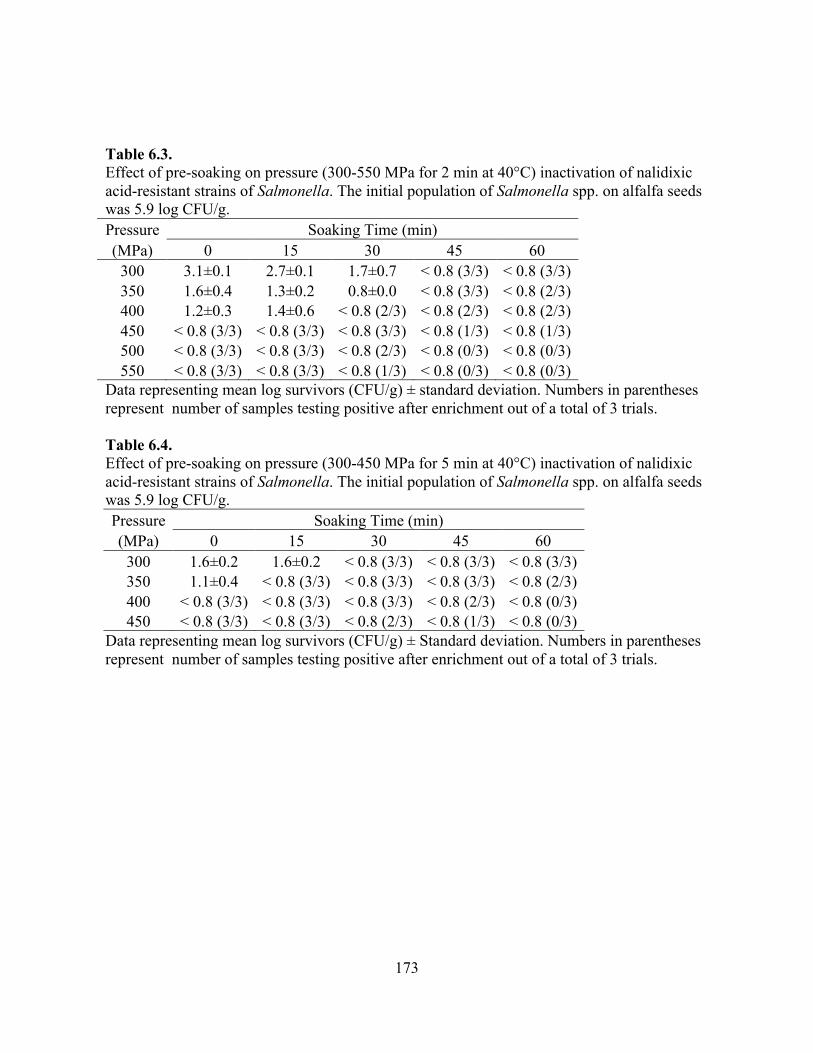
173
Table 6.3. Effect of pre-soaking on pressure (300-550 MPa for 2 min at 40°C) inactivation of nalidixic acid-resistant strains of Salmonella. The initial population of Salmonella spp. on alfalfa seeds was 5.9 log CFU/g. Pressure Soaking Time (min) (MPa) 0 15 30 45 60
300 3.1±0.1 2.7±0.1 1.7±0.7 < 0.8 (3/3) < 0.8 (3/3) 350 1.6±0.4 1.3±0.2 0.8±0.0 < 0.8 (3/3) < 0.8 (2/3) 400 1.2±0.3 1.4±0.6 < 0.8 (2/3) < 0.8 (2/3) < 0.8 (2/3) 450 < 0.8 (3/3) < 0.8 (3/3) < 0.8 (3/3) < 0.8 (1/3) < 0.8 (1/3) 500 < 0.8 (3/3) < 0.8 (3/3) < 0.8 (2/3) < 0.8 (0/3) < 0.8 (0/3) 550 < 0.8 (3/3) < 0.8 (3/3) < 0.8 (1/3) < 0.8 (0/3) < 0.8 (0/3)
Data representing mean log survivors (CFU/g) ± standard deviation. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials. Table 6.4. Effect of pre-soaking on pressure (300-450 MPa for 5 min at 40°C) inactivation of nalidixic acid-resistant strains of Salmonella. The initial population of Salmonella spp. on alfalfa seeds was 5.9 log CFU/g. Pressure Soaking Time (min) (MPa) 0 15 30 45 60
300 1.6±0.2 1.6±0.2 < 0.8 (3/3) < 0.8 (3/3) < 0.8 (3/3) 350 1.1±0.4 < 0.8 (3/3) < 0.8 (3/3) < 0.8 (3/3) < 0.8 (2/3) 400 < 0.8 (3/3) < 0.8 (3/3) < 0.8 (3/3) < 0.8 (2/3) < 0.8 (0/3) 450 < 0.8 (3/3) < 0.8 (3/3) < 0.8 (2/3) < 0.8 (1/3) < 0.8 (0/3)
Data representing mean log survivors (CFU/g) ± Standard deviation. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials.

174
Table 6.5. Effect of selected high pressure treatment conditions on seed germination rate (%).
Treatment Conditions Days of Germination Pa(MPa) Tb(°C) tc(min) Std(min) 3 4 5 6 7 8
600 40 2 0 91±1 92±1 94±1 95±2 96±2 96±2 500 45 2 0 91±2 93±0 96±2 96±2 97±3 97±3 500 40 5 0 90±1 93±1 94±2 96±1 96±1 96±1 400 45 5 0 87±3 90±2 94±1 94±1 95±1 95±1 350 50 5 0 70±4 79±5 83±5 85±5 85±5 85±5 500 40 2 45 46±1 54±1 58±2 58±2 58±2 58±2 400 40 5 60 51±8 58±7 59±7 60±7 60±7 60±7
Control 98±1 99±0 100±0 100±0 100±0 100±0 a: P = Pressure, b: T = Temperature, c: t = Pressure holding time, d: St = Soaking time before
pressure treatment
Table 6.6. Assessment of the robustness of selected treatments with respect to complete elimination of wild-type Salmonella and E. coli O157:H7. The initial populations of Salmonella spp. and E. coli O157:H7on alfalfa seeds were 5.8 and 5.2 log CFU/g, respectively.
Treatment Conditions Frequency of Survivors Pa(MPa) Tb(°C) tc(min) Salmonella E. coli O157:H7
600 40 2 (1/3) ND 500 45 2 (0/3) (0/3) 500 40 5 (0/3) (1/3)
1. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials. 2. a: P = Pressure, b: T = Temperature, c: t = Pressure holding time, ND = Not Done.

175
Chapter 7
Pre-soaking of seeds enhances pressure inactivation of E. coli O157:H7 and Salmonella
spp. on crimson clover, red clover, radish and broccoli seeds

176
Abstract
The application of high hydrostatic pressure (HHP) at a level of 600 MPa at 20°C to
decontaminate crimson clover, red clover, radish and broccoli seeds inoculated with E. coli
O157:H7 and Salmonella were evaluated. Salmonella was generally more pressure-resistant
than E. coli O157:H7 on clover and radish seeds except on broccoli seeds where the trend was
reversed. In addition, the application of HHP differentially affected seeds’ germinability and
the order of pressure tolerance of the seeds was such that red clover > crimson clover ≈
broccoli > radish seeds with final germination percentages ranging from 85-100% while their
untreated counterparts had final germination percentages of 99-100%. Pre-soaking the
different types of seeds in water for 30, 60 or 90 min at ambient temperature followed by
HHP at 600 MPa for 2 or 5 min at 20°C significantly (P < 0.05) enhanced the pressure
inactivation of the inoculated pathogens. Moreover, the ability of HHP-treated seeds to
germinate also varied as a function of the pre-soaking duration and the seed type. Pre-soaking
radish and broccoli seeds for 30 min prior to HHP (2 or 5 min) resulted in germination
percentages of ≤ 1% after 8 days of incubation. On the contrary, red clover seeds displayed
higher germination potential when pre-soaked for 60 min at 20°C prior to HPP (5 min) with
final germination percentages of 94%, although their yield was substantially lower than their
untreated counterparts. Red clover seeds pre-soaked for 60 min at 4°C followed by HPP at
600 MPa for 5 min at 20°C produced germination percentages of 91 and 95% after 3 and 8
days of sprouting compared to 99 and 100% respectively for untreated seeds. In addition, this
condition did not significantly (P > 0.05) reduce the sprout yield. The treatment also resulted
in a reduction of a 5 log initial load of E. coli O157:H7 and Salmonella to an undetectable
level (neither pathogen was detected in 2-g seed samples after enrichment).

177
Introduction
Seed sprouts are popular ingredients in many dishes and recently, they have grown in
popularity in various cultures from East to West (Wigmore, 1986). Despite being inexpensive
and easy to grow, sprouts are known to provide one of the most concentrated and naturally
occurring sources of vitamins, minerals, enzymes and amino acids known. Sprouts belonging
to the Cruciferae family (broccoli and radish sprouts) are particularly renowned for their
antioxidant and anticancerous properties (Meyerowitz, 1999a; b). Unfortunately, their rich
nutritional content also makes them very supportive of bacterial growth (Thompson and
Powell, 2000; Wood, 2000) including the growth of pathogens (Tournas, 2001; Buck et al.,
2003).
From 1973 to present, more than 41 outbreaks of foodborne illness have been reported
worldwide incriminating raw seed sprouts (International Food Safety Network, 2011).
Salmonella spp., Escherichia coli O157:H7 and Bacillus cereus were among the pathogens
involved (Fett, 2006). The most prevalent pathogenic microorganisms are enteric pathogens
such as Salmonella and E. coli O157:H7 (NACMCF, 1999). Although sprouts can become
contaminated during the sprouting, harvesting, packaging and distribution processes, many
studies demonstrated outbreaks linked to contaminated seeds (Andrews et al., 1982). Among
various seed sprouts implicated in outbreaks were alfalfa (NACMCF, 1999; Taormina et al.,
1999), radish (Taormina et al., 1999) and clover sprouts (Taormina et al., 1999; Brooks et al.,
2001).

178
The United States Food and Drug Administration thus recommends the treatment of
sprouting seeds with 20,000 ppm of free chlorine from calcium hypochlorite or an equivalent
antimicrobial (NACMCF, 1999). However, the use of chlorine-based chemicals generates
hazardous fumes that have created public health concerns (Beuchat, 1997). As a result,
alternative seed decontamination methods have been sought. Researchers studying the
efficacy of different chemical, physical and biological methods for reducing or eliminating
populations of bacterial pathogens artificially contaminated onto seeds have reported varying
degrees of success (Fett, 2006).
In our previous studies (Neetoo et al., 2008; 2009a; b), we reported that the application
of high pressure either alone or in combination with mild heat was able to successfully
inactivate E. coli O157:H7 on alfalfa seeds while maintaining seed viability. The objective of
this study was to determine the effect of pre-soaking and high pressure treatment on
inactivation of E. coli O157:H7 and Salmonella spp. inoculated on crimson clover, red clover,
radish and broccoli seeds and on their germination ability. To simplify sentence construction,
the term “elimination” was used in this paper interchangeably with the phrase “reduction of a
5 log initial load of E. coli O157:H7 or Salmonella to an undetectable level (pathogens were
not detected in 2-g seed samples after enrichment)".
Materials and methods
Effect of pressure treatment on the extent of germination of crimson clover, red clover,
radish and broccoli seeds
Unscarified radish (Raphanus sativus), broccoli (Brassica oleracea var. italica),
crimson clover (Trifolium incarnatum) and red clover (Trifolium pratense) seeds were

179
purchased from International Specialty Supply (Cookeville, TN, USA). Two grams of each
type of seed were placed in individual 3-mil-thick nylon/polyethylene pouches (Koch
Supplies, Kansas City, MO, USA). Deionized (DI) water (3 ml for crimson clover, red clover
and broccoli seeds and 4 ml for radish seeds) was added to the pouches and those pouches
were heat-sealed. Pressure treatment of samples was carried out using a high-pressure unit
with temperature control (Model Avure PT-1, Avure Technologies, Kent, WA, USA).
Pressurization was conducted at 600 MPa for 2 min at 20°C (initial seed sample temperature
prior to pressure treatment) using water as a hydrostatic medium. The temperature of the
water-bath was monitored with a K-type thermocouple. Temperature and pressure data were
recorded every 2 s (DASYTEC USA, Bedford, NH, USA). The pressure-come-up rate was
approximately 22 MPa/s. The pressure-release was almost immediate (< 4 s). Pressurization
time reported in this study does not include the pressure come-up or release times.
To determine the germination percentage of treated and un-treated seeds, seeds were
soaked in DI-water for 3 h and 100 seeds were spread evenly on layers of wet paper towels on
a plastic rack which in turn was placed into a water-filled bucket to provide a moist
environment for the seeds. The water level was maintained below the seeds’ level. The bucket
was covered loosely with a piece of plastic film to allow exchange of air between the inside
and outside of the bucket. The bucket was kept at room temperature (~21ºC) for 8 days
(suggested by the seeds provider). The seeds were visually evaluated for germination and
sprouted seeds were counted after 3, 4, 5, 6, 7 and 8 days and discarded on each day following
enumeration. The cumulative germination percentage reached on each day was then
determined as the proportion of total number of sprouted seeds to the total number of seeds.

180
Test strains and preparation of inoculum
A cocktail of five different E. coli O157:H7 strains (250, 251, Cider, 1730 and J58)
(Neetoo et al., 2008) and a cocktail of five Salmonella enterica strains (Typhimurium T43,
Typhimurium T45, Typhimurium DT 104, Enteritidis E44 and Montevideo Mo57) (Courtesy
of Dr. Rolf Joerger, University of Delaware) were used in this study. Cells of E. coli O157:H7
and Salmonella were adapted to grow in tryptic soy broth plus 0.6% yeast extract (Difco
Laboratories, Sparks, MD, USA) supplemented with nalidixic acid to a final concentration of
50 µg/ml (Fisher Scientific, Hampton, NH, USA) (TSBYE-N). Individual cultures were
grown in TSBYE-N for 16-18 hours at 35°C. Cultures were then transferred (one loopful) into
10 ml of fresh TSBYE-N and incubated at 35°C for 24 h. Equal volumes of individual
cultures were mixed to form a five-strain cocktail of Salmonella and a five-strain cocktail of
E. coli O157:H7.
Inoculation of seeds.
A hundred-fold dilution of the five-strain culture cocktails was made in sterile 0.1%
peptone water (Fisher). Five hundred grams of each seed type were added to the diluted cell
suspension at a seed:inoculum ratio of 5:3 and gently stirred for 5 min. The seeds were
separated from the cell suspension by pouring the mixture over a double layer of cheesecloth
supported by a wire screen and dried inside a biosafety hood at room temperature (21°C ±
1°C) for 48 h with intermittent rotation to ensure that seeds were uniformly dry. The mean
water activity (aw) values of radish, broccoli, crimson clover and red clover seeds after drying
were ca. 0.74, 0.44, 0.52 and 0.67, respectively. Seeds with an approximate inoculation level
of 105 CFU/g of E. coli O157:H7 and Salmonella were placed in plastic pouches and stored at

181
4°C.
Pressure treatment and microbiological analysis
Two grams of inoculated seeds mixed with 3 ml (for crimson clover, red clover and
broccoli seeds) or 4 ml (for radish seeds) of sterile DI water were added into a
nylon/polyethylene pouch. To avoid leakage during pressure treatment, each sample pouch
was placed in a larger pouch of an 8-mil-thick polyvinyl chloride plastic (McMaster-Carr,
Elmhurst, IL, USA) and heat-sealed. Pressure treatment was carried out at 20°C at 600 MPa
and a treatment time of 2, 4, 6, 8, 10 or 15 min.
Pouches containing treated seeds were cut open aseptically. The sample was
transferred into a stomacher bag to which 8 ml of sterile 0.1% peptone water was added and
subsequently stomached for 2 min at 260 rpm (Seward 400 Stomacher; Seward Medical Co.,
London, UK). The seed slurry was serially diluted in sterile 0.1% peptone water and surface
plated in duplicate on tryptic soy agar with 0.6% yeast extract (Difco) supplemented with
nalidixic acid to a final concentration of 50 µg/ml (TSAYE-N). TSAYE-N plates were
incubated for 3 days at 35°C.
Effect of the pre-soaking duration on the pressure-inactivation of E. coli O157:H7 and
Salmonella spp.
Two grams of seeds inoculated with E. coli O157:H7 or Salmonella spp. were soaked
in 20 ml sterile DI-water at room temperature (ca. 21°C) for 0 (without soaking), 30, 60, 90,
120 and 180 min. At the end of the soaking period, the excess water was subsequently
decanted and seeds were placed into a pouch in the presence of 3 ml (clover and broccoli
seeds) or 4 ml (radish seeds) of fresh sterile DI water and pressure-treated at 600 MPa for 2
min at 20°C. Samples were then microbiologically assayed on TSAYE-N as described

182
previously. When the bacterial population was below the detection limit for the plating
method (< 0.8 log CFU/g), the seed slurry was further enriched in 90 ml of TSBYE-N and
incubated for 48 h at 35°C to allow resuscitation of sub-lethally injured cells. Samples were
streaked onto Sorbitol MacConkey agar (Difco) plates supplemented with 50 µg/ml of
nalidixic acid for samples inoculated with E. coli O157:H7 or Xylose Lysine Deoxycholate
(XLD) agar (Difco) plates supplemented with 50 mg/ml of nalidixic acid for samples
inoculated with Salmonella. After 24 h incubation, presence of colorless or faint orange
growth typical of E. coli O157:H7 and black-centered or black colonies characteristic of
Salmonella were interpreted as a positive result.
In addition, the different types of seeds were also soaked for 0, 15, 30, 45 or 60 min
and then treated at 600 MPa for 5 min at 20°C. Broccoli seeds were soaked for up to 30 min
while crimson and red clover seeds were soaked for a maximum of 60 min. Treated seed
samples were then microbiologically analyzed as described above. Pre-soaked and pressure-
treated (2 min) seed samples for which no pathogens were detected in 2-g sample after
enrichment, were not further tested under the extended pressure exposure of 5 min. In
addition, treated samples without detectable pathogens after enrichment after a particular
soaking period and pressure treatment of 5 min, were not further tested under longer soaking
times.
Effect of selected soaking and pressure treatment conditions on the germination
percentages of crimson clover, red clover, radish and broccoli seeds
To determine the effect of selected soaking and pressure treatment parameters on the
seeds’ germination potential, two grams of un-inoculated seeds were soaked in 20 ml DI-
water for 30, 60 or 90 min at 4 and/or 20°C (conditions depending on the seed type). At the

183
end of the soaking period, the excess water was subsequently decanted and seeds were placed
into a pouch in the presence of 3 ml of fresh DI water (or 4 ml for radish seeds) and pressure-
treated at 600 MPa and 20°C for 2 or 5 min. Untreated (control) and pressure-treated seeds
were then soaked in DI water after pressure-treatment for a total soaking time of 3 h. One
hundred seeds were drawn from the samples and set to germinate as described previously. The
cumulative percentage of germinated seeds was determined after 3, 4, 5, 6, 7 and 8 days from
the onset of germination.
To determine whether the selected soaking conditions and pressure treatments could
eliminate the two pathogens in red clover seeds, seeds inoculated with E. coli O157:H7 and
Salmonella were (i) soaked at 4°C for 60 min and treated at 600 MPa for 5 min at 20°C or (ii)
soaked at 4°C for 90 min and treated at 600 MPa for 2 min at 20°C. Treated seeds were then
assayed post-treatment for the presence of E. coli O157:H7 or Salmonella survivors as
previously described.
Effect of selected soaking and pressure treatment conditions on the yield ratio of red
clover and crimson clover seeds
Red and crimson clover seeds (2 g) were either soaked at 4°C or 20°C for 60 min and
treated at 600 MPa for 5 min at 20°C or soaked at 4°C or 20°C for 90 min and treated at 600
MPa for 2 min at 20°C. One hundred seeds from each sample were then randomly picked and
allowed to germinate. After 8 days of growth, sprouts were weighed and the yield ratio was
calculated by dividing the weight of sprouted seeds by the weight of one hundred dry seeds, a
method adapted from Rajkowski and Thayer (2001).
Statistical analysis.
All experiments were replicated three times. Where appropriate, statistical analyses

184
were conducted using Minitab release 15 (Minitab Inc., University Park, PA, USA). One-way
analysis of variance and Tukey’s one-way multiple comparisons were used to determine
differences in the populations of E. coli O157:H7 and Salmonella recovered on treated
sprouting seeds and differences in the germination percentages and sprout yield ratio.
Differences were considered statistically significant at the 95% confidence level (P < 0.05).
Results and discussion
Effect of high pressure on the extent of germination of crimson clover, red clover,
radish, and broccoli seeds
The germination percentages for the four different types of seeds after high pressure
treatment are summarized in Table 7.1. Seed germination response was differentially affected
by high pressure and the order of pressure tolerance was such that red clover > crimson clover
≈ broccoli > radish seeds. The germination percentage determined 3-8 days after sprouting
was initially lower, but not significantly lower for red clover seeds at all germination times
and some treatment and germination times for the other seed types. Red clover seeds
exhibited the highest pressure-resistance achieving an average seed germination of 97% (after
3 days) compared with 99% for untreated seeds; showing therefore that there was almost no
delay in the onset of sprouting. Crimson clover and broccoli seeds were slightly more affected
by pressure since they sprouted to ~ 85% after 3 days and achieved > 90% germination after 4
days while control untreated seeds germinated to 99 % after 3 days. This indicates that the
onset of sprouting was slightly delayed. The germination of radish seeds appeared the most
desynchronized with an average of 14% after 3 days of germination. By the end of 8 days, the
cumulative germination percentages of red and crimson clover and broccoli seeds were 100,

185
94 and 96% respectively while radish seeds which were the most severely affected, reached a
final germination percentage of 85.
The pressure resistance of red clover seeds was very similar to those of alfalfa seeds as
investigated previously by Neetoo et al. (2008; 2009a; b). Alfalfa seeds treated at 600 MPa for
2 min at 20°C had > 95% germination after 8 days of incubation. Hayward (1948) noted that
red clover seeds and alfalfa seeds represent two very popular species of the same large pea
family, which may contribute to their similar degree of pressure resistance. The germination
results of radish seeds are also in agreement with those of Wuytack et al. (2003). They
compared the germination percentages of garden cress, mustard, radish and sesame seeds
subjected to levels of high pressure ranging from 250-400 MPa and noted that radish and
mustard seeds were highly sensitive to pressure with germination percentages < 10% after 11
days of germination.
It is not clear however what mechanism is responsible for the differential pressure
sensitivity across the various plant species studied but it is likely due to the intrinsic
uniqueness of the embryos of seeds themselves. It could also be due to the specific structural
and anatomical characteristics of the seed coat itself. Seed coat acts as an important moisture
barrier in certain seeds (Rolston, 1978) such as alfalfa, clover and broccoli seeds. Alfalfa
seeds tend to have a relatively impermeable seed coat that effectively slows down water
uptake. Lute (1928) showed that the thickened outer wall of palisade cells (and possibly the
cuticle) of alfalfa seeds constitutes a major moisture barrier. Bhalla and Slattery (1984)
demonstrated the progressive deposition of callose, a plant polysaccharide at the
plasmodesmata of clover seeds particularly in the parenchyma layer of the seed coat, thus

186
increasing the impermeability of seeds to water uptake. Wahlen (1929) also showed that the
longevity of clover seeds depended on the relative impermeability of their seed coat.
McCormac and Keefe (1990) showed that the intact seed coat (testa) of cauliflower seeds
were capable of acting as an effective barrier to water influx, thus acting as a protection
mechanism for the embryo. Dixon (2007) noted that broccoli morphologically follows similar
developmental patterns to cauliflower, hence pointing to the high degree of structural and
anatomical similarity between both types of cruciferous seeds. Hence, it is possible that
alfalfa, clover and broccoli seeds with relatively impervious coats only imbibe minimal
amount of water during the short time frame of pressurization (< 5 min); thus allowing seeds
to remain in a physiologically dormant stage. On the contrary, seeds with more permeable
seed coats readily imbibe water. Hayward (1948) described the outer surface of radish seed
coats as relatively more “pitted”, possibly rendering the seed coats more permeable to water
and hence more susceptible to the effects of high pressure. As Simon (1984) mentioned, water
uptake allows seeds to hydrate and advance into a metabolically active phase. As a result,
enzymes and other molecules critical for the development of the embryo may be activated and
more likely to be denatured under the effects of high pressure.
Pressure inactivation curve for E. coli O157:H7 and Salmonella spp. on crimson clover,
red clover, radish and broccoli seeds
The pressure inactivation curves for E. coli O157:H7 and Salmonella for each type of
sprouting seed are shown in Fig. 7.1. For both pathogens, there was a direct relationship
between the extent of bacterial inactivation and pressure exposure time; population reductions
increased with increasing holding time. After a pressure treatment time of 15 min at 600 MPa

187
at 20°C, the reductions for Salmonella spp. were 1.9, 2.1, 2.6 and 3.6 log CFU/g for crimson
clover, radish, red clover and broccoli seeds, respectively. Processing under the same
conditions achieved 2.5, 2.9, 3.0 and 2.5 log CFU/g reduction in the population of E. coli
O157:H7 on the four seeds, respectively. E. coli O157:H7 displayed significantly (P < 0.05)
higher pressure resistance than Salmonella in the case of broccoli seeds. However overall,
Salmonella spp. was more baro-tolerant than E. coli O157:H7 in crimson clover, red clover
and radish seeds although the difference was not statistically significant (P > 0.05). In the
current study, a composite of five strains of Salmonella including S. Typhimurium DT 104
were used to prepare a cocktail. This strain was included since it has become the object of
increasing concern as a result of its rapid unprecedented rapid spread through Britain and the
United States over the past 10 to 15 years (Keene, 2009). Humphrey et al. (1997) stated that
Salmonella Typhimurium DT 104 exhibited higher resistance to heat compared to other
salmonellae strains. It is possible that the salmonellae strains used in our study including S.
Typhimurium DT 104 were also particularly highly piezotolerant. As far as the substrate is
concerned, there was no significant difference (P > 0.05) in the baro-tolerance of E. coli
O157:H7 inoculated on the different sprouting seeds. Unlike E. coli O157:H7, Salmonella
was significantly (P < 0.05) more pressure sensitive on broccoli seeds compared to crimson
clover and radish seeds after a 15-min treatment.
Wuytack et al. (2003) previously pressure treated garden cress seeds inoculated with
seven different bacteria with an inoculum of 107 CFU/ml at 300 MPa for 15 min at 20°C. The
authors observed differential pressure inactivation across bacterial species ranging from 2-6
log CFU/g, with greater population reductions of gram-negative bacteria than gram-positive

188
bacteria. The authors noted that garden cress seeds, being members of the Brassicaceae or
Cruciferae family, are known to produce isothiocyanates that have inherent antimicrobial
properties (Isshiki et al., 1992; Delaquis and Massa, 1995; Lin et al., 2000 a; b). In fact, both
Wuytack et al. (2003) and Ogawa et al. (2000) agreed that gram-negative bacteria can be
sensitized to high pressure in the presence of allyl isothiocyanate (AIT). We thus speculate
that broccoli seeds, also members of the Brassicacea (or Cruciferae) family, may also be
producing isothiocyanates that may similarly sensitize gram-negative bacteria such as
Salmonella to high pressure. Cruciferous vegetables including broccoli, are rich sources of
sulfur-containing compounds called glucosinolates. Isothiocyanates are biologically active
breakdown products of glucosinolates produced upon hydrolysis by the endogenous enzyme
myrosinase. Several authors had previously identified various isothiocyanate derivatives from
broccoli using high performance liquid chromatography methods (Betz and Fox, 1994) and
the gas chromatography/mass spectrometry methods (Chiang et al., 1998; Jin et al., 1999).
Moreover, Van Eylen et al. (2007; 2009) recently showed that high pressure could induce the
conversion of glucosinolates to isothiocyanates in broccoli. This might explain the higher
reductions in the populations of Salmonella on broccoli seeds treated at 600 MPa for 15 min
at 20°C compared to the other seeds. The fact that E. coli O157:H7 displayed comparatively
higher resistance to the application of high pressure in the presence of putative
isothiocyanates present in broccoli seeds (compared to Salmonella) is corroborated by
findings of Delaquis and Sholberg (1997) who showed that the application of AIT decreased
viable S. Typhimurium to a greater extent than E. coli O157:H7.
The inactivation curves for Salmonella and E. coli O157:H7 for all tested substrates

189
exhibited a biphasic shape characterized by a rapid initial drop followed by tailing caused by a
diminishing inactivation rate. Patterson et al. (1995) reported that with the application of
hydrostatic pressure, the possibilities of surviving tail populations were more likely. It is not
uncommon to observe that a survival curve versus treatment time is concave with a rapid
initial decrease in log of survivors followed by a tailing effect, where there is essentially no
further inactivation as treatment time increases. Such inactivation curves have also been found
with other species such as Vibrio parahaemolyticus, Listeria monocytogenes and Yersinia
enterocolitica (Metrick et al., 1989; Earnshaw et al., 1995; Isaacs et al., 1995; Patterson et al.,
1995; Chen and Hoover, 2004; Chen, 2007).
Optimizing the parameters for pre-soaking seeds required for the high pressure
elimination of E. coli O157:H7 and Salmonella spp. and seed viability retention
Table 7.2 shows that treatment of 600MPa for 2 min at 20°C without prior soaking
yielded a reduction of 0.2-2.1 log CFU/g of either pathogen on the various seeds. No
significant (P > 0.05) reduction in the counts of E. coli O157:H7 and Salmonella with final
populations ranging from 4.9-5.3 log CFU/g were observed with crimson clover, red clover,
radish and broccoli seeds soaked in sterile DI water for up to 180 min without pressure
treatment (soaked controls).
However, high pressure preceded by a soaking step delivered significantly (P < 0.05)
greater inactivation of E. coli O157:H7. In fact, a direct relationship between the degree of
pressure inactivation and soaking time was observed. The same trend was noted with
Salmonella; the longer the soaking time, the greater the degree of pressure inactivation (P <
0.05). This phenomenon was possibly because the longer the time the seeds were left in

190
contact with water, the more bacteria that migrated from the inner regions of the seed to the
superficial areas of the seed coat (Charkowski et al., 2001), thus making them more
vulnerable to pressure inactivation. We speculate that longer soaking times allowed water to
permeate deeper into the cracks and crevices of the seeds thereby raising their local water
activity and hence enhancing the pressure inactivation of the cells trapped in these spaces.
The presence of E. coli O157:H7 on crimson clover seeds soaked for 60 min followed
by high pressure treatment of 2 min was undetectable after enrichment; however, survivors of
Salmonella were still detected. When the pre-soaking step was extended to 90 min followed
by a 2 min-pressure treatment, an initial load of a 5 log CFU/g Salmonella was eliminated in
the 2-g seed sample. In the case of radish, broccoli and red clover seeds, soaking for 30, 60
and 90 min respectively followed by a high-pressure treatment for 2 min was adequate for
elimination of both pathogens.
Since the length of pre-soaking negatively impacts on the seeds’ germination potential
(Neetoo et al., 2009b), we investigated whether shortening the soaking time while extending
the pressure holding time could still decontaminate seeds with similar efficacy. Results for
this study are presented in Table 7.3. Overall, an extension in the pressure exposure time at
600 MPa from 2 to 5 min reduced the soaking time requirement to 30, 60 and 60 min for
elimination of both pathogens in a 2-g sample of broccoli, crimson and red clover seeds
respectively. It is thought that a longer pressure holding time enhanced inactivation of the
pathogens as water under high pressure is forced into the deep cracks and crevices of the seed
coat allowing water to reach bacteria hidden in sub-surface locations.
Table 7.4. shows the germination percentages achieved with the various seeds soaked

191
and pressure-treated under the different conditions. Seeds soaked followed by pressure
treatment germinated to varying extents although their initial (day 3) and final (day 8)
germination percentages were lower than the untreated controls across the relevant seed types.
Un-soaked pressure-treated seeds retained their viability to a greater extent than their soaked
pressure-treated counterparts. Neetoo et al. (2009b) previously demonstrated that the
germinability of alfalfa seeds varied with soaking time; the longer the soaking duration, the
greater the severity of the treatment on the seeds. We postulate that the longer the imbibition
time in water, the greater the extent to which seeds hydrate and become physiologically active
and the more delicate and pressure-sensitive they become. Simon (1984) mentioned that seeds
are usually quiescent and can be stored for months without harm. However, once they are
supplied with water, they undergo a gain in fresh weight and embark on a second different
phase of activity marked by increased physiological and metabolic reactions as the seeds
prepare to germinate. In line with our observation, Blaszczak et al. (2007) compared the
structural and physiological changes undergone in raw and sprouted pressure-treated chickpea
seeds and observed that pressure treatment was more deleterious to germinated than raw
seeds.
In addition, we observed that seed species belonging to the Cruciferae family (radish
and broccoli) were more severely affected by high pressure than those of the Leguminosae
family (red and crimson clover). Overall, the pressure resistance of pre-soaked seeds was in
the order of red clover > crimson clover > broccoli ≈ radish seeds. We attribute this difference
principally to the characteristics of the seed coat. Barton (1961) stated that the seed coat
(testa) plays a critical role in the life and viability maintenance of seeds. We thus surmise that

192
certain seeds (red and crimson clover seeds) respond better to high pressure in the soaked
state than others (broccoli and radish seeds) by virtue of the unique characteristics of their
seed coat. Rees (1911) provided evidence that waxy (hydrophobic) cuticles common in
leguminous seeds such as alfalfa and clover seeds constitute the most impermeable part of the
seed coat in many species and the thicker the cuticle is, the longer the imbibition time
required to bring about swelling of seeds and the less severe the damage is. Wahlen (1929)
showed that in leguminous seeds such as clover seeds, their longevity was critically
dependent on the imperviousness of the seed coat. Overall, our study agrees well with our
previous findings on alfalfa seeds, alluding to the general conclusion that seeds belonging to
the Leguminosae family exhibited greater viability retention after pre-soaking and pressure
treatment than seeds belonging to the Brassicacea family. Overall, high pressure treatment on
red clover seeds soaked for 60 min and pressure-treated for 5 min produced higher initial and
final germination percentages than the 2-min pressure treatment following soaking for 90 min.
It was also the most promising treatment of all, producing a final germination percentage of
94%.
Since red and crimson clover seeds produced more satisfactory results after soaking
and high pressure treatment compared to radish and broccoli seeds with respect to their
germinability, we subsequently determined whether the extent of germination could be
enhanced by lowering the pre-soaking temperature. Results of viability tests for red and
crimson clover seeds pre-soaked for 60 or 90 min at refrigeration temperature prior to
pressure treatment at 600 MPa for 5 and 2 min, respectively are included in Table 7.4. Pre-
soaking at a lower temperature (4°C) significantly (P < 0.05) enhanced the germinability of

193
both clover seed types regardless of the pre-soaking duration. As Barton (1961) described, in
general, the higher the temperature, the more rapid is the rate of deterioration of the seeds’
germinability at a given moisture level. Conversely, the lower the temperature, the greater the
seeds’ tolerance for high moisture content. Barton (1961) also mentioned that at a low to
moderate moisture content, maintaining the temperature in the range of 5-10°C, will extend
the life of seeds beyond that achieved under similar humidity conditions at ordinary room
temperature. Overall, the treatments appeared to be more feasible for red clover seeds than
crimson clover seeds as the former seed type achieved final germination percentages of 95
and 88% after pre-soaking for 60 and 90 min respectively compared to 75 and 56% for the
latter. Moreover, soaking for 60 or 90 min at 4°C followed by HHP did not significantly (P >
0.05) affect the yield ratio of red clover seeds (14.3 and 12.1, respectively) compared to
control untreated seeds (15.0). However, all other treatment conditions significantly (P <
0.05) reduced the yield ratio of red and crimson clover seeds as shown in Table 7.4.
We then finally evaluated the efficacy of low temperature soaking of red clover seeds
for 60 and 90 min followed by HPP at 600 MPa for 5 and 2 min respectively to decontaminate
red clover seeds challenged with E. coli O157:H7 and Salmonella. Pre-soaking for 60 min at
4°C followed by high-pressure treatment for 5 min was able to eliminate both pathogens;
while pre-soaking for 90 min followed by a pressure exposure of 2 min resulted in detectable
E. coli O157:H7 survivors in one out of three trials.
Conclusions
The results of our study demonstrated that the application of high hydrostatic pressure
inactivated E. coli O157:H7 and Salmonella on various leguminous and cruciferous seeds. We

194
also showed that a pre-soaking step carried out at ambient temperature considerably enhanced
pressure inactivation of either enteric pathogen with variable effects on the seed
germinability. High-pressure treatment on soaked seeds appeared more promising for
leguminous than cruciferous seeds. Finally, we showed that pre-soaking seeds at refrigeration
temperature followed by high pressure treatment decontaminated red clover seeds, achieving
a final germination percentage of 95% with minimal impact on the sprout yield.
Acknowledgement
This study was supported by a start-up fund from the Department of Animal and Food
Sciences at the University of Delaware. We wish to thank Dr. Thompson Pizzolato at the
University of Delaware for the helpful discussion.

195
References Andrews, W.H., Mislivec, P.B., Wilson, C.R., Bruce, V.R., Poelma, P.L., Gibson, R., 1982. Microbial hazards associated with bean sprouting. Journal of the Association of Official Analytical Chemists 65, 241-248. Barton, L.V., 1961. Seed preservation and longevity. Interscience Publishers, Inc., New York, U.S.A., pp. 14-28, 30-39 Betz, J.M., Fox, W.D., 1994. High-performance liquid chromatographic determination of glucosinolates in Brassica vegetables. ACS symposium series 546, 181-196. Beuchat, L.R., 1997. Comparison of chemical treatments to kill Salmonella on alfalfa seeds destined for sprout production. International Journal of Food Microbiology 34, 329-333. Bhalla, P.L., Slattery, H.D., 1984. Callose deposits make clover seeds impermeable to water. Annals of Botany 53, 125-128. Blaszczak, W., Doblado, R., Frias, J., Vidal Valverde, C., Sadowska, J., Fornal, J., 2007. Microstructural and biochemical changes in raw and germinated cowpea seeds upon high-pressure treatment. Food Research International 40, 415–423. Brooks, J.T., Rowe, S.Y., Shillam, P., 2001. Salmonella Typhimurium infections transmitted by chlorine-pretreated clover sprout seeds. American Journal of Epidemiology 154, 1020–1028 Buck, J.W., Walcott, R.R., Beuchat, L.R., 2003. Recent trends in microbiological safety of fruits and vegetables. Plant Health Progress. <http:// www.plantmanagementnetwork.org> Accessed August 2009. Charkowski, A.O., Sarreal, C.Z., Mandrell, R.E., 2001. Wrinkled alfalfa seeds harbor more aerobic bacteria and are more difficult to sanitize than smooth seeds. Journal of Food Protection 64, 1292-1298. Chen, H.Q., Hoover, D.G., 2004. Use of Weibull model to describe and predict pressure inactivation of Listeria monocytogenes Scott A in whole milk. Innovative Food Science and Emerging Technologies 5, 269-276. Chen, H., 2007. Use of linear, Weibull, and log-logistic functions to model pressure inactivation of seven foodborne pathogens in milk. Food Microbiology 24, 197-204.

196
Chiang, W.C.K., Pusateri, D.J., Leitz, R.E.A., 1998. Gas chromatography/mass spectrometry method for the determination of sulforaphane and sulforaphane nitrile in broccoli. Journal of agricultural and food chemistry 46, 1018-1021. Delaquis, P.J., Massa, G., 1995. Antimicrobial properties of isothiocyanates in food preservation. Food Technology 11, 73-84. Delaquis, P.J., Sholberg, P.L., 1997. Antimicrobial activity of gaseous allyl isothiocyanate. Journal of Food Protection 60, 943-947. Dixon, G.R., 2007. Vegetable brassicas and related crucifers. Crop Production Science in Horticulture Series, CABI Publisher, Cambridge, MA, pp. 91-113. Earnshaw, R.G., Appleyard, J., Hurst, R.M., 1995. Understanding physical inactivation processes: combined preservation opportunities using heat, ultrasound and pressure. International Journal of Food Microbiology 28, 197-219. Fett, W.F., 2006. Interventions to ensure the microbial safety of sprouts. In: Sapers, G.M., Gorny, J.R., Yousef, A.E. (Eds.), Microbiology of Fruits and Vegetables. Taylor and Francis Group, LLC, Boca Raton, FL., pp. 187-210. Hayward, H.E., 1948. The structure of economic plants. Macmillian Company, Chicago, U.S.A., pp. 309-339. Humphrey, T.J., Wilde, S.J., Rowbury, R.J., 1997. Heat tolerance of Salmonella Typhimurium DT104 isolates attached to muscle tissue. Letters in Applied Microbiology 25, 265-268. Isaacs, N.S., Chilton, P., Mackey, B., 1995. Studies on the inactivation by pressure of micro-organisms. In: Ledward, D.A., Johnston, D.E., Earnshaw, R.G., Hasting, A.P.M. (Eds.), High Pressure Processing of Foods. Nottingham University Press, Nottingham, U.K., pp. 65-79. Isshiki, K., Tokuora, K., Mori, R., Chiba, S., 1992. Preliminary examination of allyl isothiocyanante vapor for food preservation. Bioscience, Biotechnology and Biochemistry 56, 1476-1477. Jin, Y., Wang, M., Rosen, R.T., Ho, C.T., 1999. Thermal degradation of sulforaphane in aqueous solution. Journal of Agricultural and Food Chemistry 47, 3121-3123. Keene, W.E., 1999. Lessons from investigations of foodborne disease outbreaks. Journal of American Medical Association 281, 1845-1847. Lin, C.M., Kim, J., Du, W., Wei, C., 2000. Bactericidal activity of isothiocynate against pathogens on fresh produce. Journal of Food Protection 63, 25-30.

197
Lin, C.M., Preston, J.F., Wei, C., 2000. Antibacterial mechanism of ally isothiocyanate. Journal of Food Protection 65, 383-388. Lute, A.M., 1928. Impermeable seed of alfalfa. Colorado Agricultural Experimental Station Bulletin, Volume 326. McCormac, A.C., Keefe, P.D., 1990. Cauliflower (Brassica oleracea L.) seed vigour: imbibition effects. Journal of Experimental Botany 41, 893-899. Metrick, C., Hoover, D.G., Farkas, D.F., 1989. Effects of high hydrostatic pressure on heat-resistant and heat-sensitive strains of Salmonella. Journal of Food Science 54, 1547-1549. Meyerowitz, S., 1999a. Sprouts: the miracle food. Sproutman Publications, Massachusetts, U.S.A., pp. 57-79. Meyerowitz, S., 1999b. My favorite greens. Better nutrition 61, 18-21.
NACMCF, 1999. Microbiological safety evaluations and recommendations on sprouted seeds. International Journal of Food Microbiology 52, 123-153. Neetoo, H., Ye, M., Chen, H., 2008. Potential application of high hydrostatic pressure to eliminate Escherichia coli O157:H7 on alfalfa sprouted seeds. International Journal of Food Microbiology 128, 348-353. Neetoo, H., Pizzolato, T., Chen, H., 2009a. Elimination of Escherichia coli O157:H7 from alfalfa seeds through a combination of high hydrostatic pressure and mild heat. Applied Environmental Microbiology 75, 1901-1907. Neetoo, H., Ye, M., Chen, H., 2009b. Factors affecting the efficacy of pressure inactivation of Escherichia coli O157:H7 on alfalfa seeds and seed viability. International Journal for Food Microbiology 131, 218-23. Ogawa, T., Nakatani, A., Matsuzaki, H., Isobe, S., Isshiki, K., 2000. Combined effects of hydrostatic pressure, temperature and addition of allyl isothiocynanate on inactivation of Escherichia coli. Journal of Food Protection 63, 884-888. Patterson, M.F., Quinn, M., Simpson, R., Gilmour, A., 1995. Effects of high pressure on vegetative pathogens. In: Ledward, D.A., Johnston, D.E., Earnshaw, R.G., Hasting, A.P.M. (Eds.), High Pressure Processing of Foods. Nottingham University Press, pp.47-63. Rajkowski, K.T., Thayer, D.W., 2001. Alfalfa seed germination and yield ratio and alfalfa sprout microbial keeping quality following irradiation of seeds and sprouts. Journal of Food

198
Protection 64, 1988-1995. Rees, B., 1911. Longevity of seeds and structure and nature of seed coats. Proceedings of the Royal Society of Victoria 23, 393-414. Rolston, P., 1978. Water impermeable seed dormancy. Botanical Review 44, 365-389. Simon, E.W., 1984. Early events in germination. In: Murray, D.R. (Ed.), Seed Physiology, Germination and reserve mobilization. Academic Press, Australia 2, pp. 77-115. Taormina, P.J., Beuchat, L.R., Slutsker, L., 1999. Infections associated with eating seed sprouts: an international concern. Emerging Infectious Diseases 5, 626-634. Thompson, S., Powell, D.A., 2000. Risks associated with the consumption of fresh sprouts, Food Safety Network Technical Report # 16, http://www.plant.uoguelph.ca/safe-food/micro-haz/sprouts-risk-sylvanus-juloo.htm
Tournas, V.H., 2001. Moulds and yeasts in fresh and minimally processed vegetables, and sprouts. International Journal of Food Microbiology, 99, 618–622. Van Eylen, D., Oey, I., Hendrickx, M., Van Loey, A., 2007. Kinetics of the stability of broccoli (Brassica oleracea cv. Italica) myrosinase and isothiocyanates in broccoli juice during pressure/temperature treatments. Journal of Agricultural and Food Chemistry 55, 2163-2170.
Van Eylen, D., Bellostas, N., Strobel, B.W., Oey, I., Hendrickx, M., Van Loey, A., Sørensen, H., Sørensen, J.C., 2009. Influence of pressure/temperature treatments on glucosinolate conversion in broccoli (Brassica oleraceae L. cv Italica) heads. Food Chemistry 112, 646-653. Wahlen, F.T., 1929. Hard seededness and longevity in clover seeds. Proceedings of the International Seed Test Association 9-10, 34-39. Waje, C.K., Han, D.H., Jo, C., Kwon, J.H., Jun, S.Y., Lee, Y.K., Kim, B.N., 2009. Microbial quality assessment and pathogen inactivation by electron beam and gamma irradiation of commercial seed sprouts. Food Control 20, 200-204. Wigmore, A. 1986. The Sprouting Book. Avery, Penguin Group, U.S.A., pp. 5-12. Wood, M., 2000. Safer sprouts. Agricultural research, Washington, D.C., 48, 16-17. Wuytack, E.Y., Michiels, C.W., Meersseman, K., Diels, A.M., 2003. Decontamination of seeds for seed sprout production by high hydrostatic pressure. Journal of Food Protection 66, 18-923.
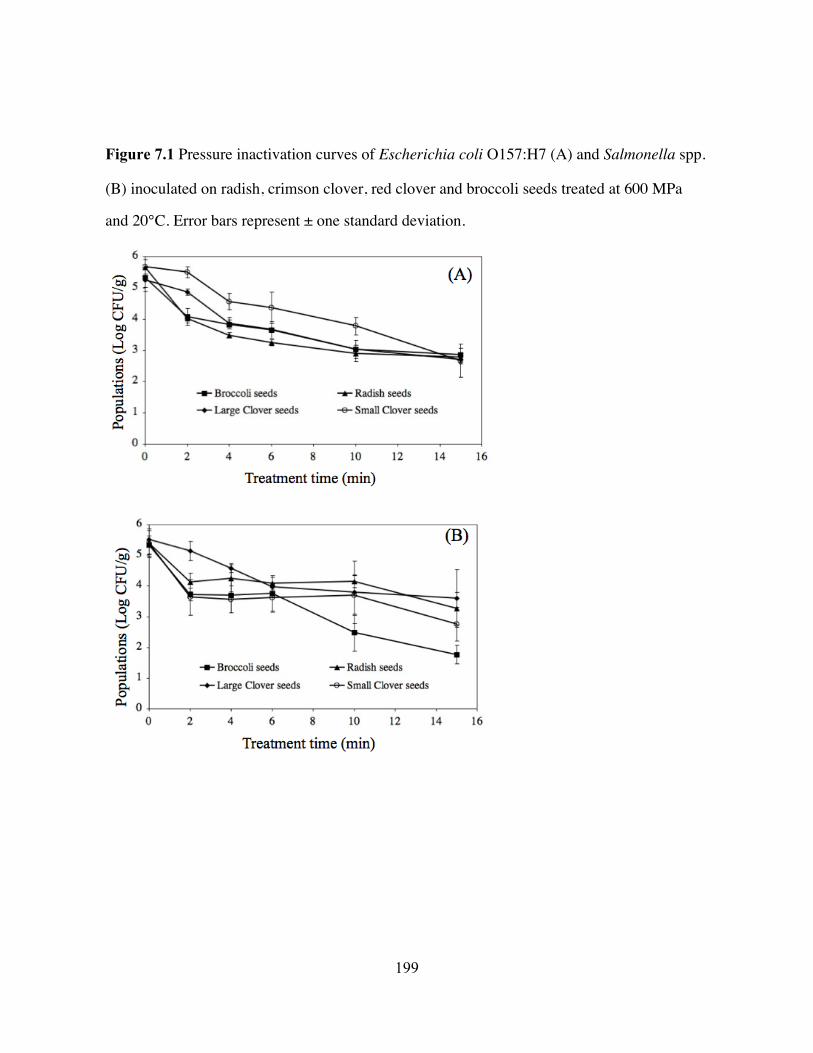
199
Figure 7.1 Pressure inactivation curves of Escherichia coli O157:H7 (A) and Salmonella spp.
(B) inoculated on radish, crimson clover, red clover and broccoli seeds treated at 600 MPa
and 20°C. Error bars represent ± one standard deviation.
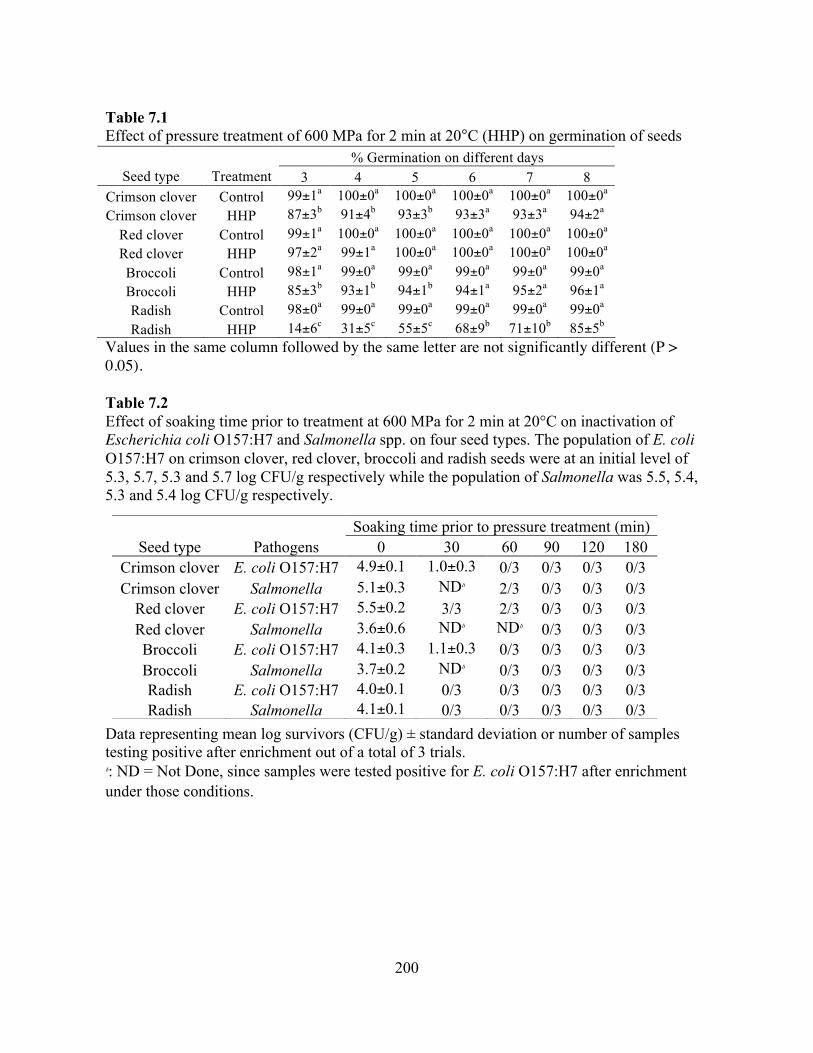
200
Table 7.1 Effect of pressure treatment of 600 MPa for 2 min at 20°C (HHP) on germination of seeds
% Germination on different days Seed type
Treatment 3 4 5 6 7 8
Crimson clover Control 99±1a 100±0a 100±0a 100±0a 100±0a 100±0a Crimson clover HHP 87±3b 91±4b 93±3b 93±3a 93±3a 94±2a
Red clover Control 99±1a 100±0a 100±0a 100±0a 100±0a 100±0a Red clover HHP 97±2a 99±1a 100±0a 100±0a 100±0a 100±0a Broccoli Control 98±1a 99±0a 99±0a 99±0a 99±0a 99±0a Broccoli HHP 85±3b 93±1b 94±1b 94±1a 95±2a 96±1a Radish Control 98±0a 99±0a 99±0a 99±0a 99±0a 99±0a Radish HHP 14±6c 31±5c 55±5c 68±9b 71±10b 85±5b
Values in the same column followed by the same letter are not significantly different (P > 0.05). Table 7.2 Effect of soaking time prior to treatment at 600 MPa for 2 min at 20°C on inactivation of Escherichia coli O157:H7 and Salmonella spp. on four seed types. The population of E. coli O157:H7 on crimson clover, red clover, broccoli and radish seeds were at an initial level of 5.3, 5.7, 5.3 and 5.7 log CFU/g respectively while the population of Salmonella was 5.5, 5.4, 5.3 and 5.4 log CFU/g respectively.
Data representing mean log survivors (CFU/g) ± standard deviation or number of samples testing positive after enrichment out of a total of 3 trials. δ: ND = Not Done, since samples were tested positive for E. coli O157:H7 after enrichment under those conditions.
Soaking time prior to pressure treatment (min) Seed type Pathogens 0 30 60 90 120 180
Crimson clover E. coli O157:H7 4.9±0.1 1.0±0.3 0/3 0/3 0/3 0/3 Crimson clover Salmonella 5.1±0.3 NDδ 2/3 0/3 0/3 0/3
Red clover E. coli O157:H7 5.5±0.2 3/3 2/3 0/3 0/3 0/3 Red clover Salmonella 3.6±0.6 NDδ NDδ 0/3 0/3 0/3 Broccoli E. coli O157:H7 4.1±0.3 1.1±0.3 0/3 0/3 0/3 0/3 Broccoli Salmonella 3.7±0.2 NDδ 0/3 0/3 0/3 0/3 Radish E. coli O157:H7 4.0±0.1 0/3 0/3 0/3 0/3 0/3 Radish Salmonella 4.1±0.1 0/3 0/3 0/3 0/3 0/3

201
Table 7.3 Effect of soaking time prior to treatment at 600 MPa for 5 min at 20°C on the inactivation of E. coli O157:H7 and Salmonella spp. on four sprouting seed types. The population of E. coli O157:H7 on crimson clover, red clover, broccoli and radish seeds were at an initial level of 5.5, 5.2, 5.3 and 5.4 log CFU/g respectively while the population of Salmonella was 5.5, 5.2, 5.3 and 5.7 log CFU/g respectively. Soaking time prior to pressure treatment (min)
Seed type Pathogens 0 15 30 45 60 Crimson clover E. coli O157:H7 3.8±0.1 1.7±0.3 2/3 0/3 0/3† Crimson clover Salmonella 4.4±0.3 NDδ NDδ 2/3 0/3
Red clover E. coli O157:H7 4.5±0.4 2/3 2/3 1/3 0/3 Red clover Salmonella 3.6±0.6 NDδ NDδ NDδ 0/3 Broccoli E. coli O157:H7 3.7±0.2 1.7±0.3 0/3 Broccoli Salmonella 3.8±0.2 NDδ 0/3 Radish E. coli O157:H7 3.4±0.1 3/3 0/3† Radish Salmonella 4.1±0.3 NDδ 0/3†
Data representing mean log survivors (CFU/g) ± standard deviation or number of samples testing positive after enrichment out of a total of 3 trials. δ: ND = Not Done, since samples were tested positive for E. coli O157:H7 after enrichment under those conditions. †: Inferred from data of Table 7.2 and/or Table 7.3 rather than experimentally determined.
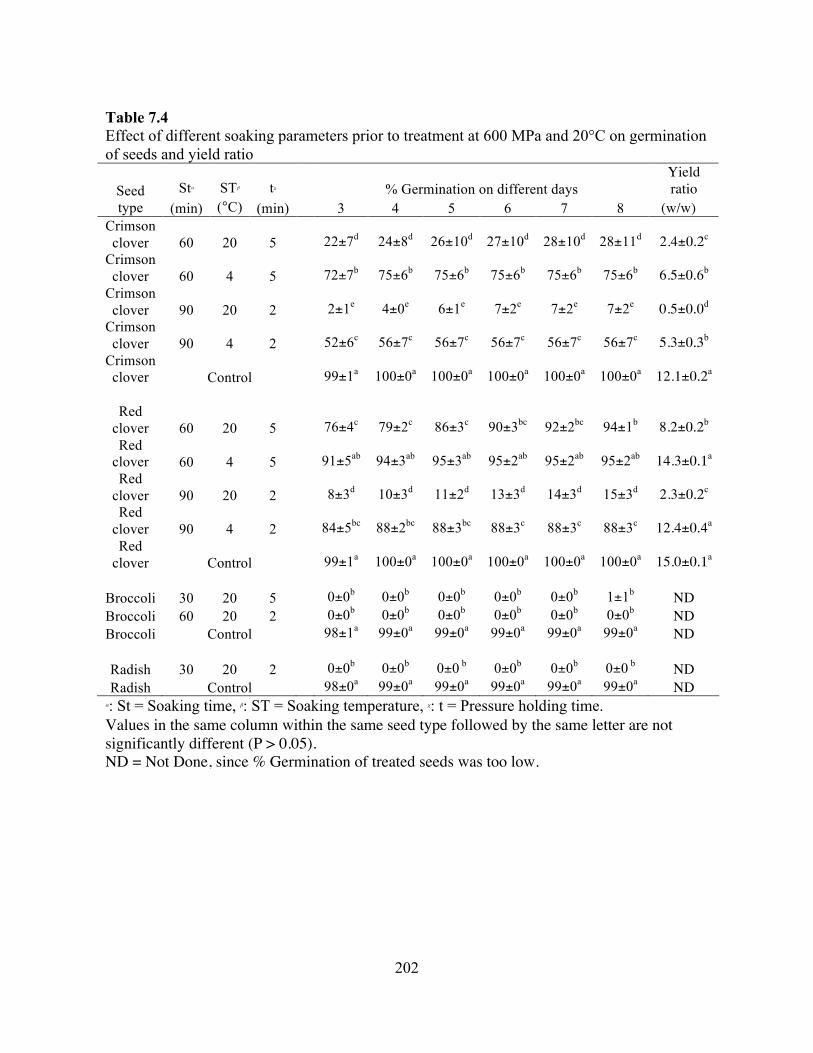
202
Table 7.4 Effect of different soaking parameters prior to treatment at 600 MPa and 20°C on germination of seeds and yield ratio
Stα STβ tχ % Germination on different days Yield ratio Seed
type (min) (°C) (min) 3 4 5 6 7 8 (w/w) Crimson clover 60 20 5 22±7d 24±8d 26±10d 27±10d 28±10d 28±11d 2.4±0.2c
Crimson clover 60 4 5 72±7b 75±6b 75±6b 75±6b 75±6b 75±6b 6.5±0.6b
Crimson clover 90 20 2 2±1e 4±0e 6±1e 7±2e 7±2e 7±2e 0.5±0.0d
Crimson clover 90 4 2 52±6c 56±7c 56±7c 56±7c 56±7c 56±7c 5.3±0.3b
Crimson clover Control 99±1a 100±0a 100±0a 100±0a 100±0a 100±0a 12.1±0.2a
Red
clover 60 20 5 76±4c 79±2c 86±3c 90±3bc 92±2bc 94±1b 8.2±0.2b Red
clover 60 4 5 91±5ab 94±3ab 95±3ab 95±2ab 95±2ab 95±2ab 14.3±0.1a Red
clover 90 20 2 8±3d 10±3d 11±2d 13±3d 14±3d 15±3d 2.3±0.2c Red
clover 90 4 2 84±5bc 88±2bc 88±3bc 88±3c 88±3c 88±3c 12.4±0.4a Red
clover Control 99±1a 100±0a 100±0a 100±0a 100±0a 100±0a 15.0±0.1a
Broccoli 30 20 5 0±0b 0±0b 0±0b 0±0b 0±0b 1±1b ND Broccoli 60 20 2 0±0b 0±0b 0±0b 0±0b 0±0b 0±0b ND Broccoli Control 98±1a 99±0a 99±0a 99±0a 99±0a 99±0a ND
Radish 30 20 2 0±0b 0±0b 0±0 b 0±0b 0±0b 0±0 b ND Radish Control 98±0a 99±0a 99±0a 99±0a 99±0a 99±0a ND
α: St = Soaking time, β: ST = Soaking temperature, χ: t = Pressure holding time. Values in the same column within the same seed type followed by the same letter are not significantly different (P > 0.05). ND = Not Done, since % Germination of treated seeds was too low.

203
Chapter 8
Individual and combined application of dry heat with high hydrostatic pressure to
inactivate Salmonella and Escherichia coli O157:H7 on alfalfa seeds
Published I n Food Microbiol.

204
Abstract
Alfalfa sprouts are recurrently implicated in outbreaks of food-borne illnesses as a
result of contamination with Salmonella or Escherichia coli O157:H7. In the majority of these
outbreaks, the seeds themselves have been shown to be the most likely source of
contamination. The aims of this study were to comparatively assess the efficacy of dry heat
treatments alone or in conjunction with high hydrostatic pressure (HHP) to eliminate a ~5 log
CFU/g load of Salmonella and E. coli O157:H7 on alfalfa seeds. Dry heat treatments at mild
temperatures of 55 and 60°C achieved ≤ 1.6 and 2.2 log CFU/g reduction in the population of
Salmonella spp. after a 10-d treatment, respectively. However, subjecting alfalfa seeds to
more aggressive temperatures of 65°C for 10 days or 70°C for 24 h eliminated a ~ 5 log
population of Salmonella and E. coli O157:H7. We subsequently showed that the sequential
application of dry heating followed by HHP could substantially reduce the dry heating
exposure time while achieving equivalent decontamination results. Dry heating at 55, 60, 65
and 70°C for 96, 24, 12 and 6 h, respectively followed by a pressure treatment of 600 MPa for
2 min at 35°C were able to eliminate a ~ 5 log CFU/g initial population of both pathogens.
Finally, we evaluated the impact of selected treatments on the seed germination percentages
and yield ratios and showed that dry heating at 65°C for 10 days did not bring about any
considerable decrease in the germination percentage. However, the sprout yield of treated
alfalfa seeds was reduced by 21%. Dry heating at 60 and 65°C for 24 and 12 h respectively
followed by the pressure treatment of 600 MPa for 2 min at 35°C did not significantly (P >
0.05) affect the germination percentage of alfalfa seeds although a reduction in the sprouting
yield was observed.

205
Introduction
Seed sprouts represent an important component of traditional oriental cuisine by virtue
of their excellent nutritional profiles, and recently have gained increased popularity in many
parts of the world, including Europe and the United States. Sprouts are believed by many to
have anti-cholesterolemic, anti-carcinogenic and anti-oxidative activities (Meyerowitz, 1999).
Unfortunately, the increased consumption of sprouts throughout the world has led to a
concomitant surge in the incidence of food-borne illnesses associated with this commodity
(Fett & Cooke, 2005; Fett et al., 2005).
Salmonella ranks as one of the most frequently reported cause of food-borne outbreaks
of gastroenteritis in the United States, making salmonellosis an increasing public health
concern (CDC, 2008). In the United States alone, there have been at least 29 reports of sprout-
borne Salmonella outbreaks, involving more than 1500 cases since 1995 (Taormina et al.,
1999; Chapman, 2009), with most outbreaks incriminating alfalfa sprouts. However, the
safety of sprouts has also become an international concern with the occurrence of outbreaks in
Canada (Van Beneden et al., 1999), the United Kingdom (O’ Mahony et al., 1990), Japan
(Gutierrez, 1997) and several European countries including Finland, Sweden (Ponka et al.,
1995; Puohiniemi et al., 1997) and Denmark, (Van Beneden et al., 1999). Salmonella has
been more frequently identified as the etiological agent involved in sprout-related illnesses
compared to other food-borne pathogens such as Escherichia coli O157:H7 and Bacillus
cereus (Proctor et al., 2001; Hora et al., 2007).
Based on the reported isolation of pathogens from implicated seed lots or on the basis
of epidemiological evidence, seeds harboring pathogenic bacteria are the most likely source of

206
contamination of sprouts (Fett et al., 2005). Although sprouting seeds represent very low
moisture foods with an average water activity (aw) of ca. 0.3, contaminated seeds still present
a safety concern since it is widely known that Salmonella can persist for extended periods of
time in low moisture foods (Betts, 2007; Zink, 2007). As a result, a few pathogenic cells on
seeds can multiply to potentially hazardous levels because of the favorable conditions of
moisture, temperature and nutrient availability during subsequent seed germination and sprout
growth.
In response to the urgent need to mitigate the occurrence of outbreaks, the U.S. Food
and Drug Administration and the California Department of Health Services enforce that
growers treat sprouting seeds with an approved antimicrobial treatment such as 20,000 ppm of
free chlorine from calcium hypochlorite immediately prior to sprouting (Fett et al., 2005). To
date, this treatment still remains the yardstick against which the efficacy of other aqueous
sanitizers is compared. However, the limited performance of calcium hypochlorite along with
the fact that organic sprout growers are not allowed to treat seeds with such a high level of
chlorinated sanitizer, has spurred a search for alternative seed decontamination treatments. A
variety of physical treatments have been tested as stand-alone interventions for
decontamination of sprouting seeds in the form of thermal or non-thermal interventions.
Thermal treatments investigated have included hot water, dry heat, radio-frequency dielectric
heating while non-thermal technologies have included gamma irradiation, hydrostatic
pressure, pulsed UV light and ultrasound.
Thermal treatments used for the decontamination of vegetative materials, including
seeds, have been one of the most heavily studied intervention methods and has a long history

207
of use since the 1920s. Hot water treatments for reducing populations of human pathogens on
sprout seeds were first studied by Jaquette et al. (1996). They found that a 5 min-treatment in
hot water at 57 to 60°C led to a 2.5 log unit decrease in the population of S. enterica on
artificially contaminated seeds with no appreciable reduction in the seed germination rate.
However, temperatures that were slightly higher or heating durations that were slightly longer
brought about significant decrease in seed germinability. However, the application of dry heat
treatment to decontaminate mung bean and alfalfa seeds has been shown to yield more
encouraging results (Beuchat & Scouten, 2002; Hu et al., 2004; Feng et al., 2007).
In our previous studies, we had demonstrated that the application of high pressure at
650 MPa for 15 min at 20°C completely eliminated a ~5 log CFU/g initial burden of E. coli
O157:H7 on alfalfa seeds (Neetoo et al., 2008). We subsequently showed that the application
of high hydrostatic pressure at a reduced level of 300-600 MPa for 2 or 5 min in combination
with mild heat at temperatures ranging from 40-50°C, was able to successfully eliminate E.
coli O157:H7 (Neetoo et al., 2009) and Salmonella (Neetoo & Chen, 2010) on alfalfa seeds
while maintaining seed viability. While the aforementioned treatments did not unacceptably
decrease the percent germination, a variable reduction in the lengths of the sprouts was
observed. The study reported here was undertaken to compare the influence of dry heat alone
and the combined action of dry heat and HHP on the lethality to Salmonella and E. coli
O157:H7 on alfalfa seeds and seed viability. To simplify sentence construction, the term
“elimination” will be used in this paper with the intended meaning of “reduction of a 5 log
initial load of E. coli O157:H7 or Salmonella to an undetectable level in 2-g seed samples
after enrichment".

208
Materials and methods
Bacterial Strains
Five Salmonella and five E. coli O157:H7 strains were used in this study. The
Salmonella strains included two Salmonella enterica serovar Typhimurium strains (T43 and
TDT 104), Salmonella enterica serovar Enteritidis E44, Salmonella enterica serovar
Montevideo Mo57 and Salmonella enterica serovar Seftenberg 775W. The E. coli O157:H7
strains included strain 1730, Cider strain (Ibrahim et al., 2006), strains 250 and 251 (Bhagwat
et al., 2005) and J58 (Barak et al., 2005). All the strains were from the University of Delaware
culture collection. The cells of Salmonella or E. coli O157:H7 were adapted to grow in tryptic
soy broth plus 0.6% yeast extract (Difco Laboratories, Sparks, MD) supplemented with
nalidixic acid to a final concentration of 50 µg/ml (Fisher Scientific, Hampton, NH) (TSBYE-
N). Individual cultures were grown in TSBYE-N overnight at 35°C. Cultures were then
transferred (one loopful) into 10 ml of fresh TSBYE-N and incubated at 35°C for 24 h. Equal
volumes of individual cultures of Salmonella or E. coli O157:H7 were mixed to form a five-
strain composite.
Inoculation of seeds
A hundred-fold dilution of the five-strain culture cocktails was made in sterile 0.1%
peptone water (Fisher). Five hundred grams of alfalfa seeds were added to 300 ml of peptone
water containing the diluted cell suspension (~ 7 log CFU/ml) and mixed well on a magnetic
stirrer for 5 min. The seeds were separated from the cell suspension by pouring the mixture
over a double layer of cheesecloth supported by a wire screen and dried inside a biosafety
hood at room temperature (21 ± 2°C) for up to 72 h with intermittent rotation to ensure that

209
seeds were uniformly dry. The inoculated seeds were placed in plastic pouches and stored at
4°C for at least 3 days prior to subjecting them to any treatment. Seeds inoculated with
Salmonella and E. coli O157:H7 were used within 4 and 2 weeks of inoculation respectively,
since a preliminary study indicated that the populations of Salmonella and E. coli O157:H7
were stable (< 0.5 log CFU/g decline) during storage at 4°C for up to 30 and 15 days,
respectively. The initial microbial counts of both Salmonella and E. coli O157:H7 were also
determined right before treatments and were at an approximate level of 5 log CFU/g of
Salmonella or E. coli O157:H7 as reported in the results.
Dry heat treatment
Inoculated and un-inoculated seeds (2g) were placed in capped glass tubes and
subjected to dry heating at temperatures ranging from 55-70°C (in 5°C increments). Samples
of seeds were heated at 55, 60 and 65°C for up to 10 days and 70°C for up to 24 h. After heat-
treatment of seeds at 55 and 60°C, seeds were allowed to equilibrate to ambient temperature
(22-23°C) for 1 h prior to microbiological analysis (for inoculated seeds) and seed viability
test (for un-inoculated seeds). Seeds treated at 65 and 70°C were refrigerated overnight at 4°C
prior to microbiological analysis (for inoculated seeds) and seed viability test (for un-
inoculated seeds). These post-treatment storage conditions were chosen in the light of
preliminary experiments showing that seeds dry heated at 55 and 60°C and microbiologically
analyzed immediately after treatment had lower bacterial counts than if seeds were allowed to
“rest” for at least 1 h (data not shown). No appreciable difference in the microbial counts was
observed for plating done 1 h or 24 h post-treatment. In addition, we did not observe any
difference in the counts of treated seeds kept at ambient or refrigeration temperature. For

210
seeds treated at 65 and 70°C on the contrary, we observed that a 24-h repair phase at 4°C was
optimum for recovery of heat-stressed cells (data not shown) and thus adequate for providing
an accurate estimate of numbers of survivors.
Microbiological analysis
Seeds were transferred into a stomacher bag to which 8 ml of sterile 0.1% peptone
water was added and subsequently stomached for 2 min at 260 rpm (Seward 400 Stomacher;
Seward Medical Co., London, United Kingdom). The seed mixture was serially diluted in
sterile 0.1% peptone water and surface-plated in duplicate on tryptic soy agar with 0.6% yeast
extract (Difco Laboratories, Sparks, MD) supplemented with nalidixic acid to a final
concentration of 50 µg/ml (TSAYE-N). TSAYE-N plates were incubated for 3 days at 35°C.
Presumptive colonies of Salmonella or E. coli O157:H7 formed on the plates were
enumerated. In addition, the seed mixture was also directly enriched in 90 ml of TSBYE-N
and incubated for 48 h at 35°C to allow resuscitation of sub-lethally injured cells where
appropriate. Samples were streaked onto Sorbitol MacConkey agar (Difco) plates
supplemented with 50 µg/ml of nalidixic acid for samples inoculated with E. coli O157:H7 or
Xylose Lysine Deoxycholate (XLD) agar (Difco) plates supplemented with 50 mg/ml of
nalidixic acid for samples inoculated with Salmonella. After 24 h incubation, presence of
colorless or faint orange growth typical of E. coli O157:H7 and black-centered or black
colonies characteristic of Salmonella were interpreted as a positive result.
Determination of seed germination percentage
Un-inoculated seeds were dry-heated at temperatures of 55-70°C as described above.
To determine the seed germination percentage, heat-treated and control (non heat-treated) un-

211
inoculated seeds were soaked in DI-water for 3 h. One hundred seeds were then randomly
picked and dispersed evenly on layers of wet paper towels laid on a plastic rack, which in turn
was placed into a water-filled bucket to provide a moist environment for the seeds. The water
level was maintained below the seeds’ level. The bucket was covered loosely with a piece of
plastic film to allow exchange of air with its surroundings. The bucket was kept at room
temperature (21 ± 2ºC) for 8 days (suggested by the seeds provider). The seeds were visually
evaluated for germination and sprouted seeds were counted after 3, 4, 5, 6, 7 and 8 days and
discarded. The germination percentage was determined as the proportion of sprouted seeds to
the total number of seeds. The cumulative germination percentage on each day was then
computed.
Combined effects of dry heat and high-pressure treatment on the inactivation of
Salmonella on alfalfa seeds
The sequential application of dry heat and high hydrostatic pressure (HHP) to
inactivate Salmonella was subsequently investigated. Inoculated alfalfa seeds were dry-heat
treated (2 g) at 55, 60, 65 and 70°C to varying durations depending on the temperature. After
the dry heat treatment, seeds dry-heated at 55 and 60°C were allowed to equilibrate to room
temperature for 1 h while seeds heated at 65 and 70°C were refrigerated overnight. Seeds
were then subsequently placed in individual 3-mil thick nylon/polyethylene pouches (Koch
Supplies, Kansas City, MO, USA). Sterile de-ionized (DI) water (3 ml) was then added to the
pouches, which were heat-sealed. Pressure treatment of samples was carried out using a high-
pressure unit with temperature control (Model Avure PT-1, Avure Technologies, Kent, WA,
USA) with pressurization conducted at 600 MPa for 2 min at 20 or 35°C (initial seed sample

212
temperature prior to pressure treatment) using water as a hydrostatic medium. The
temperature of the water-bath was monitored with a K-type thermocouple. Temperature and
pressure data were recorded every 2 s (DASYTEC USA, Bedford, NH, USA). The pressure-
come-up rate was approximately 22 MPa/s. The pressure-release was almost immediate (< 4
s). Pressurization time reported in this study does not include the pressure come-up or release
times.
For samples to be pressure-treated at 35°C, samples were first submerged in the 35°C
water bath surrounding the pressure cell for 10 min, allowing samples to equilibrate to the
water bath temperature before pressurization. The temperature equilibration step and the
actual pressure treatment constituted a brief seed-soaking period lasting ca. 15 min.
Immediately after pressure treatment, samples were cooled in an ice-water mixture. Samples
were then microbiologically assayed as described above.
Assessment of the efficacy of selected treatments to achieve ~ 5 log reduction of
E. coli O157:H7 to below undetectable levels
Two grams of seeds inoculated with an initial burden of ~ 5 log CFU/g of E. coli
O157:H7 were dry heated at 55°C for 96 h, 60°C for 24 h, 65°C for 12 h and 70°C for 6 h.
After dry heat treatment, seeds were allowed to rest at room or refrigeration temperature as
described previously before being mixed with 3 ml of DI water, packaged, and treated at 600
MPa for 2 min at 35°C. Two grams of alfalfa seeds inoculated with E. coli O157:H7 were
also subjected to dry heat at 65°C for 10 d and 70°C for 24 h. Samples were then
microbiologically assayed on TSAYE-N or enriched as described previously.

213
Determination of the germination percentages and yield ratio of treated seeds
To determine the possible impact of various decontamination treatments on the seeds’
germination potential, 2 g of un-inoculated alfalfa seeds were subjected to dry heat treatment
at 65°C for 10 days or dry heat in combination with high pressure treatment. For the
combined treatment, 2 g of seeds was heat treated at 55°C for 96 h, 60°C for 24 h, 65°C for
12 h or 70°C for 6 h. Following dry heating, these seeds were allowed to rest at room or
refrigeration temperature as mentioned before and then mixed with 3 ml of DI water,
packaged, and treated at 600 MPa for 2 min at 35°C. After pressure-treatment, seeds were
immediately chilled in iced water for 10 min and then subsequently soaked in water for 3 h.
One hundred seeds from each treated sample were then randomly picked and allowed to
germinate using the procedure described above. Germinated seeds were counted after 3-8
days of germination. In addition, sprouts were weighed after 8 days of growth and the yield
ratio was calculated by dividing the weight of sprouted seeds by the weight of one hundred
dry seeds, a method adapted from Rajkowski & Thayer (2001).
Modification of the germination procedure to improve the sprouting yield of seeds
treated by dry heat and HHP
Un-inoculated seeds were dry heated at 60°C for 24 h and 65°C for 12h as described
previously. Following dry heating, seeds were kept at 4°C for 24 h and then pressure-treated
at 600 MPa for 2 min at 35°C. Seeds were then chilled, soaked and allowed to germinate for
3-8 days at 27°C. We used 27°C as the incubation temperature of choice since we observed
slightly higher germination rates at 27°C compared to 22°C and 30°C (data not shown).

214
Germinated seeds were enumerated after 3-8 days of germination. In addition, sprouted seeds
were weighed after eight days and the sprout yield determined as described above.
Statistical analysis
All experiments were replicated at least three times. Where appropriate, statistical
analyses were conducted using Minitab® Release 15 (Minitab Inc., University Park, PA,
USA). One-way analysis of variance (ANOVA) and Tukey’s one-way multiple comparisons
were used to determine differences in the populations of Salmonella and E. coli O157:H7
recovered on treated alfalfa seeds as well as differences in the germination percentage of
seeds. Significant differences were considered at the 95% confidence level (P < 0.05).
Results and discussion
Effect of dry heat treatment on the population of Salmonella and E. coli O157:H7 on
inoculated alfalfa seeds
Tables 8.1-8.4 indicate that the level of Salmonella decreased to varying extents and at
varying rates during storage at 55, 60, 65 and 70°C respectively. At 55 and 60°C, the
population initially declined to approximately 3.7 and 3.9 log CFU/g respectively after 2 days
and remained fairly stable at a level of ca. 3.0 log CFU/g for the remaining days of the 10-d
period. However, when the temperature was increased to 65°C, we observed that there was a
steady decline in the population from an initial of ~ 5 log CFU/g to undetectable levels after
10 days. At a higher temperature of 70°C, we observed a faster decline in the population of
Salmonella to below the detection limit (< 0.7 log CFU/g) after a 24-h exposure. Hence,
exposures of seeds to high temperatures of 65 and 70°C brought about a faster decline in the
population of Salmonella compared to temperatures of 55 and 60°C which reduced the

215
populations by < 2 log CFU/g after 10 days of exposure. An extended holding period of up to
30 days at 55 and 60°C did not further reduce the population of Salmonella with final
population of ca. 3.5 log CFU/g (data not shown). Beuchat and Scouten (2002) also showed
that seeds exposed to mild temperatures of 50 and 60°C underwent a marginal reduction in
the population of Salmonella and E. coli O157:H7 of < 1.5 log CFU/g after 7 h of exposure
while heating at higher temperatures of 70 and 80°C achieved > 3 log CFU/g reduction.
Feng et al. (2007) and Hu et al. (2004), on the other hand, both reported on the
relatively higher efficacy of dry heat treatments at 55°C on mung bean and alfalfa seeds,
respectively. Hu et al. (2004) showed that seeds inoculated with cocktails of E. coli O157:H7
(6 log CFU/g) or S. enterica (4 log CFU/g) subsequently exposed to a temperature of 55°C,
resulted in undetectable levels of the pathogen after 4 and 5 days respectively. Feng et al.
(2007) showed that an exposure lasting 8 and 6 days at 55°C were able to completely
eliminate an initial population of 8 log CFU/g of Salmonella spp. and E. coli O157:H7,
respectively. It is possible that the cause of the disparity in our results lies in the variation in
the water activity of inoculated seeds or differences in the heat resistance of the strains
studied. In our study, a composite of five strains of Salmonella were used, including S.
Typhimurium DT 104 and S. Seftenberg 775W. S. Typhimurium DT 104 is known to exhibit
higher tolerance to heat and acid that many other salmonellae and has also been shown to
mount stress responses to conditions of low stresses (Humphrey, 2001). S. Seftenberg 775W
has also been reported to be fairly resistant to thermal treatments (Ng et al., 1969), with 10-20
times higher thermal resistance than typical Salmonella strains. Although several authors have
commented that this strain exhibits highest resistance at water activity close to 1.00 (Goepfert

216
& Biggie, 1968; Riemann, 1968), S. Seftenberg 775W is known to be generally quite
refractory to heat (Baird-Parker et al., 1970). These two thermally resistant strains were not
included in the cocktails used in studies undertaken by Feng et al. (2007) and Hu et al. (2004).
Doyle and Mazzotta (2000) also acknowledged that the heat resistance data acquired by
different researchers for different serovars of Salmonella enterica in low moisture food
products such as alfalfa seeds are quite variable, emphasizing that the heat resistance of
Salmonella is highly influenced by the strain(s) tested. In recognition of the differential heat
tolerance of strains, it is thus preferable to use strain cocktails in inactivation and validation
studies conducted on food (Balasubramaniam et al., 2004), and to include highly heat resistant
strains to simulate a worst-case scenario. These would be especially valuable for food
processors and may help them in designing acceptance limits on critical points that ensure
safety against the pathogen on alfalfa seeds.
Moreover, the heat resistance of salmonellae may also be influenced by the
inoculation and subsequent drying method of the seeds. In our study, alfalfa seeds were
inoculated by mixing them into the inoculum at a ratio of 5:3. The excess liquid was allowed
to drain and the seeds were allowed to dry with intermittent turn over for a total period of 72
h. The mean water activity for inoculated seeds dried for 24, 48 and 72 h were 0.703, 0.584
and 0.439 respectively. Hu et al. (2004) and Feng et al. (2007), on the contrary, only allowed
the seeds to dry at room temperature overnight. It is possible that our inoculation method and
subsequent prolonged drying of seeds for 72 h prior to the actual treatment caused the
bacterial cells to become conditioned or “habituated” at a reduced water activity, resulting in a
concomitant increase in their thermal resistance (Mattick et al., 2000). As it is well known, the

217
water activity during heat inactivation exerts a profound effect on the heat tolerance of
bacteria (Sumner et al., 1991). Kirby and Davies (1990) had previously demonstrated that air-
dried Salmonella cells acquired greater heat tolerance in a matrix of reduced aw. Mattick et al.
(2000) previously reported that transient conditioning of a bacterial cell to a sub-lethal stress
(i.e. “habituation”) can confer greater resistance to a more extreme form of the stress. Mattick
et al. (2000) further demonstrated that habituation of S. Typhimurium DT 104 to low aw at
ambient temperature (~21°C), can greatly increase its heat tolerance and the impact of
habituation on heat tolerance decreases at higher treatment temperatures suggesting the
involvement of additional targets for cell death. Observations made by the aforementioned
author(s) may thus help to explain the relatively higher thermal resistance exhibited by
Salmonella at 55 and 60°C compared to temperatures of 65 and 70°C.
Decline in the population of E. coli O157:H7 on inoculated seeds was noticeably
faster than Salmonella at all temperatures (Tables 8.1-8.4). Feng et al. (2007) and Hu et al.
(2004) also observed that E. coli O157:H7 was much more heat sensitive to dry heat than
Salmonella artificially contaminated onto alfalfa and mung bean seeds, respectively. We
speculate that this is probably because of the higher heat tolerance exhibited by one or more
strains of Salmonella, such as S. Typhimurium DT 104 and S. Seftenberg 775W. Moreover,
Barak et al. (2002) reported that Salmonella has the ability to attach more tightly to seeds and
sprout tissue than E. coli O157:H7. Recent findings by Barak et al. (2002) also suggested that
S. enterica may have a survival advantage over E. coli O157:H7 for attachment to sprouting
seeds, sprouts and plant tissue in general. The authors explained that the difference could
particularly be explained by the differential ability of S. enterica and E. coli O157:H7 to

218
produce aggregative fimbriae (curli). It is thought that S. enterica most readily express curli
(Tafi), a structure that plays a fundamental role in the attachment of S. enterica to seeds and
plant tissue in general. Romling et al. (1998) showed that the production of curli was induced
in an environment akin to a plant surface as well as low temperature and osmolarity. It is
possible that the higher thermo-tolerance of Salmonella may be attributed to its stronger
attachment to the seed tissue. Cotterill & Glauert (1971) also showed that the heat resistance
of salmonellae was much higher than microorganisms belonging to other genera in low-
moisture foods. The heat resistance of S. Typhimurium, in particular, has been shown to be
about 653 times as great in low moisture foods such as dried egg white (moisture of about
7%) compared to liquid eggs (Liu et al., 1969). Hansen and Riemann (1963) pointed out that
dry heat appears to be less efficient than moist heat in inactivating microorganisms because
proteins that are potentially destroyed during (moist) thermal processing remain quite stable
in the dry state.
Effect of dry heat treatment on seed germination percentages
With regard to the viability of seeds, alfalfa seeds exposed to dry heat temperatures of
55-65°C retained their ability to germinate to a larger extent than when held at 70°C (Tables
1-4).
Indeed, Farooq et al. (2004) reported that the effect of dry-heat treatment on seed germination
depended on both the dry-heat intensity and duration of exposure. Martin and Cushwa (1966)
also made a general observation that under a range of optimal heat treatment temperatures,
seeds normally exhibit a consistently high germination rate but result in decreased
germinability at higher lethal temperatures. Farooq et al. (2004) attributed the decreased

219
germinability at elevated detrimental temperatures to membrane deterioration. We speculate
that the rapid decline in germination rate for seeds treated at 70°C to be due to reduced
membrane integrity (Basavarajappa et al., 1991; Farooq et al., 2004) and possibly to heat-
induced damage to plastids, mitochondria or other cellular organelles as proposed by Wang
and Berjak (2000).
Extended exposures of alfalfa seeds at 55-65°C did not substantially impact on the
seed germinability with a germination percentage of > 90% after 10 days of holding at 55, 60
and 65°C. Feng et al. (2007) also showed that when seeds were exposed to a temperature of
55°C for 4 days or longer, the germination extent of treated un-inoculated seeds (78%) were
not dramatically lower compared to control untreated seeds (82%). However, the same
authors mentioned that temperatures higher than 55°C were detrimental to seed germination
although they did not specify the temperatures or temperature ranges studied. Beuchat and
Scouten (2002) did not observe any major difference in the germination percentage between
treated (81.6 %) and untreated seeds (86.3 %) at 50°C for 24 h. They also showed that the
germinability of seeds was fairly unaffected even after exposure to a higher temperature of
60°C for 3 h (87.6 %) compared to control untreated seeds (85.0 %), although the same author
reported that heating of seeds at 70°C for ≥ 3 h or at 80°C for ≥1 h, resulted in a decrease in
germination rate. This observation and the above comment made by Feng et al. (2007)
reconcile with our findings, showing that exposures to 55, 60 and 65°C had minimal adverse
effects on the seed germinability while a higher temperature of 70°C brought about a
noticeable reduction in the germination percentage to ~ 31% with dry heating times ≥ 9 h.

220
It is not surprising that alfalfa seeds are quite refractory to dry heat. As a matter of
fact, Stewart (1926) previously indicated that alfalfa seeds heated at temperatures ranging
from 60-75°C for a maximum of 1-2 h displayed higher or comparable germination rates (76-
93%) to unheated seeds (79%). They however showed that seeds underwent a drastic decrease
in viability (36%) when exposed to a brief (10 min) exposure at 80°C. The author reported
that alfalfa seeds could be heated for 1 h at 75°C without injuring the seed. Staker (1925)
found that the killing point of alfalfa seeds was about 90°C and that temperatures as high as
85°C resulted in an increase in the percentage germination.
Hence, the above findings garnered by previous researches shows that there is a
certain degree of variability among different studies. However, these data are unanimously
supportive of the fact that mild temperatures of 55 and 60°C and slightly higher temperature
of 65°C do not result in a dramatic loss in seed viability.
As Beuchat and Scouten (2002) inferred from their study, there appears to be a
correlation between loss of seed viability and loss of Salmonella viability. This is also evident
in our study by comparing the bacterial population reduction and seed germination
percentages in Tables 8.1-8.4. While temperatures of 55 and 60°C appeared to be more
‘friendly’ to seeds with minimal adverse impact on seed germination, the effectiveness of
these temperatures at reducing the population of Salmonella was also comparatively lower.
On the other hand, dry heating at 70°C caused a rapid and steep decline in pathogen
inactivation with an accompanying rapid decrease in seed viability.

221
Combined effect of dry heat and high-pressure treatment on the inactivation of
Salmonella on alfalfa seeds
When seeds with an initial burden of ~ 5 log CFU/g of Salmonella were subjected to a
dry heat treatment at 55°C for 2 days followed by pressure treatment of 600 MPa for 2 min at
20°C, the population decreased by ca. 4.3 log CFU/g (Table 8.5). Extending the dry heating
time to 4 days followed by the pressure treatment led to a final population of < 0.8 log CFU/g
(below the detection limit) although survivors were detectable after enrichment in one out of
three cases. However, when the initial temperature of the pressure treatment was stepped up
to 35°C following 4 days of dry heating at 55°C, the population was consistently undetectable
after enrichment.
When the dry heat temperature was increased to 60°C, we observed that a dry heat
treatment of 1 day followed by the pressure treatment of 600 MPa for 2 min at 20°C was able
to reduce the population to below the detection limit although survivors were still detected in
one out of three trials (Table 8.5). An increase in the initial temperature of the high pressure
processing step from 20°C to 35°C completely eliminated the population with no detectable
survivors after enrichment. Pressure-treatment of (600 MPa for 2 min at 35°C) seed samples
producing no detectable pathogens after 1 day of dry heating at 60°C were not further tested
under longer dry heating times.
Similar to the combination treatments of dry heat at 55 and 60°C with HHP, we
observed from Table 8.5 that the sequential application of dry heat at 65°C for 12 h and 70°C
for 6 h followed by the pressure treatment of 600 MPa for 2 min at 35°C was also able to
completely eliminate Salmonella from alfalfa seeds. The results from Tables 5 and 6 show

222
that the pressure sensitivity of Salmonella had a strong dependence on the HHP treatment
temperature, with increased sensitivity at a higher temperature of 35°C. Indeed, it is well
documented that initial temperatures above 30°C greatly enhance the pressure inactivation of
bacteria as reported for Listeria monocytogenes (Chen, 2007), Staphylococcus aureus (Chen
et al., 2006), Escherichia coli (Ponce et al., 1998), Salmonella enterica serovar Enteritidis
(Ponce et al., 1999), Vibrio vulnificus (Kural & Chen, 2008), and Vibrio parahaemolyticus
(Kural et al., 2008). Studies conducted by Kingsley et al. (2006) and Chen et al. (2005)
demonstrated that temperatures of 30°C also facilitated the pressure inactivation of hepatitis A
virus and feline calicivirus, respectively.
The efficacy of the conjunct application of dry heat with other hurdles has also been
investigated by other authors (Bari et al., 2003; 2009). Previous research has indicated that
dry heat treatment (50°C for 1 h) followed by an exposure to gamma irradiation (2 to 2.5
kGy) eliminated E. coli O157:H7 from artificially inoculated alfalfa, radish and mung bean
seeds (4 to 5 log CFU/g reductions) without decreasing seed germination percentages (Bari et
al., 2003; 2009). However, one of the problems encountered with gamma irradiation is the
phenomenon of uneven or patchy absorbance of irradiation at different locations in the
treatment chamber (Fett et al., 2005). As a result, the dose absorbed by seeds located near the
exterior of the chamber may be different from the dose absorbed by seeds placed at the center
of the chamber (Fett et al., 2005), thus rendering optimization and scale-up of the process
difficult. This problem of heterogeneity could lead to variability in pathogen population
reduction within a batch of treated seeds as well as possible disparate effects on the
subsequent seed viability. High pressure processing on the contrary, is governed by the

223
‘Isostatic Principle’, which states that pressure is uniformly transmitted throughout a sample
under pressure independent of the batch size (Hogan et al., 2005). Hence, the application of
dry heat in combination with high pressure would be expected to yield more consistent results
with minimal batch-to-batch variation as opposed to other gradient-forming processes such as
heat and radiation.
Assessment of the efficacy of selected treatments to achieve complete elimination of
E. coli O157:H7
Table 8.6 summarizes the effects of different dry heat time/temperature regimes alone
or in conjunction with high pressure at reducing the population of E. coli O157:H7 to
undetectable levels. Results presented showed that complete elimination of E. coli O157:H7
was possible when alfalfa seeds were heated for 10 days at 65°C and under selected
conditions of dry heat exposure time (6 h – 4 days), dry heat temperature (55-70°C) and HHP
(600 MPa for 2 min at 35°C). Based on the results presented in Tables 1-4, we expected the
strains of E. coli O157:H7 to display higher thermal sensitivity than those of Salmonella. In
addition, on the basis of our previously developed pressure inactivation curve at 600 MPa for
Salmonella (Neetoo & Chen, 2010) and E. coli O157:H7 (Neetoo et al., 2008) on alfalfa
seeds, we found that Salmonella was more baro-tolerant than E. coli O157:H7. As anticipated,
the pre-determined treatments were able to eliminate E. coli O157:H7 as well. The findings
of this study thus indicate that dry heat alone or in conjunction with high pressure can
represent effective interventions for the decontamination of alfalfa seeds from both enteric
pathogens.

224
Determination of the germination percentages and yield ratios of seeds treated under
selected conditions
The germination percentages as determined 3-8 days from the onset of sprouting
showed that seeds pressure-treated under the different conditions were affected to variable
extents depending on the conditions of the treatment (Table 8.7). Application of dry heat for
10 days at 65°C appeared as the most promising treatment with respect to seed germination
percentage (95%) and yield (12.2 w/w) compared to untreated seeds, with a germination
percentage of 100% and yield of 15.5 w/w (Table 8.7). This represents an approximate 21%
reduction in yield ratio, which may still be considered acceptable by the commercial sprout
grower (Rajkowski & Thayer, 2001).
The sequential application of dry heat at 70°C and HHP was deleterious to seed
viability with a significantly (P < 0.05) reduced germination percentage and yield. Although
seeds dry heated at 60 and 65°C and treated by HHP did not undergo an appreciable decrease
in their germination rates, we observed a significant decrease (P < 0.05) in the yield with final
sprouting yield ratios of ~ 9 w/w. Other authors have also observed that while certain seed
treatment regimes may have little to no effect on seed germination, the impact on the
sprouting yield could be considerable. Rajkowski and Thayer (2001) previously showed that
the application of gamma irradiation at doses ranging from 1-5 kGy had little apparent effect
on seed germination although the yield ratio of alfalfa sprouts decreased with increasing dose.
Bari et al. (2003) previously showed that the sequential application of dry heat at 50°C for 1 h
followed by irradiation at doses of up to 2.0 kGy had little to no effect on the germination rate
of radish and mung bean seeds although a significant decrease in sprout lengths were

225
observed. Bari et al. (2009) also recently showed that irradiation of mung bean seeds at 1.0
kGy subsequent to dry heating at 50°C for 17 h did not affect the percentage germination
although the author reported a significant decrease in the average sprout length. We can thus
ascertain that sprout yield ratio is an important index of seed viability in addition to the
percentage germination.
Modification of the germination procedure to improve the sprouting yield of pressure-
treated seeds
Previously, we showed that the viability of seeds that were dry heated at 60 and 65°C
prior to HHP was adversely affected, resulting in an appreciable decrease in their yield. In this
experiment (Table 8.7), we showed that when seeds were dry heated at 60°C for 24 h, chilled
at 4°C for 24 h prior to HHP, and subsequently allowed to germinate at an incubation
temperature of 27°C (rather than ambient temperature,) a sprouting yield of 11.4 w/w was
recorded, compared to the yield of 9.2 w/w determined previously. When seeds were dry
heated at 65°C for 12 h, chilled at 4°C for 24 h prior to HHP and then allowed to germinate
under the same conditions, only a slight improvement in the yield was observed with a final
sprouting yield ratio of 10.9 w/w compared to 9.2 w/w recorded previously. Hence,
modification to the germination conditions only brought about a marginal improvement in the
yield ratio. It is likely that although certain seed decontamination regimes may have minimal
adverse effect on the processes leading to radicle emergence (as measured by percentage
germination), seedling growth (as measured by sprout yield) appears to be more variably
impaired. Future studies are thus warranted to determine whether seeds subjected to dry
heating and high pressure could undergo a restoration of viability upon storage.

226
Conclusions
The results of this study indicate that dry heating alone at moderate temperatures (55
and 60°C) represented an ineffective intervention to decontaminate alfalfa seeds from
Salmonella. However, exposure to a relatively more aggressive temperature of 65°C with a
holding time of 10 days, was able to simultaneously eliminate Salmonella as well as E. coli
O157:H7 at little expense of the seed germinability. The study also stressed on various
intrinsic and extrinsic factors impacting on the dry heat inactivation including the choice and
type of strains used, the aw of the matrix of interest and the treatment temperatures. This
should be of particular interest to seed companies or sprout growers wishing to implement a
dry heat treatment step. In addition to a stand-alone intervention, we also demonstrated that a
‘multiple-hurdle’ approach consisting of a dry heating step (60°C/24h) in combination with
high pressure (600 MPa/ 2 min/ 35°C) fully decontaminated alfalfa seeds but also led to a >
20% reduction in the sprout yield.
Acknowledgements
This study was supported by a start-up fund from the Department of Animal and Food
Sciences at the University of Delaware.

227
References
Baird-Parker, A.C., Boothroyd, M., Jones, E., 1970. The effect of water activity on the heat resistance of heat sensitive and heat resistant strains of Salmonella. J. Appl. Bacteriol. 33, 515–522. Balasubramaniam, V.M., Ting, E.Y., Stewart, C.M., Robbins, J.A., 2004. Recommended laboratory practices for conducting high-pressure microbial experiments. Innov. Food Sci. Emerg, Technol. 5, 299-306. Barak, J.D., Whitehand, L.C., Charkowski, A.O., 2002. Differences in attachment of Salmonella Enterica serovars and Escherichia coli O157:H7 to alfalfa sprouts. Appl. Environ. Microbiol. 68, 4758-4763. Barak, J.D., Delwiche, M.J. Sananikone, K., 2005. Comparison of primers for the detection of pathogenic Escherichia coli using real-time PCR. Lett. Appl. Microbiol. 41, 112-118. Bari, M.L., Nazuka, E., Sabina, Y., Todoriki, S., Isshiki, K., 2003. Chemical and irradiation treatments for killing Escherichia coli O157:H7 on alfalfa, radish, and mung bean seeds. J. Food Prot. 66, 767-774. Bari, M.L., Todoriki, S., Kawamoto, S., Nei, D., Enomoto, K., 2009. Combination treatments for killing Escherichia coli O157:H7 on alfalfa, radish, broccoli, and mung bean seeds. J. Food Prot. 72, 631-636. Bari, M.L., Kawamoto, S., Sugiyama, J., 2009. Repeated quick hot-and-chilling treatments for the inactivation of Escherichia coli O157:H7 in mung bean and radish seeds. Foodborne Pathog. Dis. 6, 137-143. Basavarajappa, B.S., Shetty, S.H., Prakash, H.S., 1991. Membrane deterioration and other biochemical changes associated with accelerated ageing of maize seeds. Seed Sci. Technol. 19, 279–286. Betts, R., 2007. Water, water, everywhere nor any drop to drink – The problem of Salmonella in low-moisture foods. IAFP Special Interest Session on Salmonella growth, persistence and survival in low-moisture foods and their environment – strategies for control. IAFP annual meeting, July 8-11, Buena Vista, Florida. Beuchat, L.R., Scouten, A.J., 2002. Combined effects of water activity, temperature and

228
chemical treatments on the survival of Salmonella and Escherichia coli O157:H7 on alfalfa seeds. J. Appl. Microbiol. 92, 382-395. Bhagwat, A.A., Chan, L., Han, R., Tan, J., Kothary, M., Jean-Gilles, J., Tall, B.D., 2005. Characterization of enterohemorrhagic Escherichia coli strains based on acid resistance phenotypes. Infect. Immun. 73, 4993–5003. [CDC] Centers for Disease Control and Prevention. Annual listing of foodborne disease outbreaks. Available at http://www.cdc.gov/foodborneoutbraks/outbreak_data.htm.Atlanta, GA; CDC, 2008. Chapman, B., 2009. Sprouts associated outbreaks in North America, 1990-2009. Accessed on from Ben Chapman Bites, Food Safety Network. http://foodsafety.k-state.edu/en/article-details.php?a_2&c_6&sc_36&id_865. (accessed 05.14.09) Chen, H., Hoover, D.G., Kingsley, D.H., 2005. Temperature and treatment time influence high hydrostatic pressure inactivation of feline calicivirus, a norovirus surrogate. J. Food Prot. 68, 2389-2394. Chen, H., Guan, D., Hoover, D.G., 2006. Sensitivities of food-borne pathogens to pressure changes. J. Food Prot. 69, 130-136. Chen, H., 2007. Temperature-assisted pressure inactivation of Listeria monocytogenes in turkey breast meat. Int. J. of Food Microbiol. 117, 55-60. Cotterill, O.J., Glauert, J., 1971. Thermal resistance of salmonellae in egg yolk containing 10% sugar or salt after storage at various temperatures. Poult. Sci. 50, 109-115. Doyle, M.E., Mazzotta, A.S., 2000. Review of studies on the thermal resistance of salmonellae. J. Food Prot. 63, 779-795. Farooq, M., Basra, S.M.A., Hafeez, K., Warriach, E.A., 2004. Influence of high- and low-temperature treatments on seed germination and seedling vigor of coarse and fine rice. Int. Rice Res. Notes. 29, 75-77. Feng, G., Churey, J.J., Worobo, R.W., 2007. Thermal Inactivation of Salmonella and Escherichia coli O157:H7 on alfalfa seeds. J. Food Prot. 70, 1698-1703. Fett, W.F., Cooke, P.H., 2005. A survey of native microbial aggregates on alfalfa, clover and mung bean sprout cotyledons for thickness as determined by confocal scanning laser microscopy. Food Microbiol. 22, 253-259.

229
Fett, W.F., Tortorello, M., Fu, T., 2005. Seed sprouts: the state of microbiological safety. In: Matthews, K.R. (Ed.), Microbiology of Fresh Produce. ASM Press, Washington, D.C., pp. 167-219. Goepfert, J.M., Biggie, R.A., 1968. Heat resistance of Salmonella Typhimurium and Salmonella Senftenberg 775W in milk chocolate. Appl. Microbiol. 16, 1939–1940. Gutierrez, E., 1997. Japan prepares as O157 strikes again. Lancet, 349, 1156. Hansen, N.H., Riemann, H., 1963. Factors affecting the heat resistance of non-sporing organisms. J. Appl. Bacteriol. 26, 314–333. Hogan, E., Kelly, A., Sun, D.W., 2005. High pressure processing of foods: an overview. In: D.W. Sun (Ed.), Emerging technologies for food processing, Elsevier Academic Press, London, pp. 3-32. Hora, R., Kumar, M., Kostrzynska, M., Dixon, M.A., Warriner, K., 2007. Inactivation of Escherichia coli O157:H7 and Salmonella on artificially or naturally contaminated mung beans (Vigna radiata L) using a stabilized oxychloro-based sanitizer. Lett. Appl. Microbiol. 44, 188-193. Hu, H., Churey, J.J., Worobo, R.W., 2004. Heat treatments to enhance the safety of mung bean seeds. J. Food Prot. 67, 1257-1260. Humphrey, T., 2001. Salmonella Typhimurium definitive type 104: a multi-resistant Salmonella. Int. J. Food Microbiol. 67, 173-186. Ibrahim, S.A., Salameh, M.M., Phetsomphou, S., Yang, H., Seo, C.W., 2004. Application of caffeine, 1,3,7-trimethylxanthine, to control Escherichia coli O157:H7. Food Chem. 99, 645–650. Jaquette, C.B., Beuchat, L.R., Mahon, B.E., 1996. Efficacy of chlorine and heat treatment in killing Salmonella Stanley inoculated onto alfalfa seeds and growth and survival of the pathogen during sprouting and storage. Appl. Environ. Microbiol. 62, 2212-2215. Kingsley, D.H., Guan, D., Hoover, D.G., Chen, H., 2006. Inactivation of hepatitis A virus by high pressure processing: the role of temperature and pressure oscillation. J. Food Prot. 69, 2454-2459. Kirby, R.M., Davies, R., 1990. Survival of dehydrated cells of Salmonella Typhimurium LT2 at high temperatures. J. Appl. Bacteriol. 68, 241-246.

230
Kural, A.G., Chen, H., 2008. Conditions for a 5-log reduction of Vibrio vulnificus in oysters through high hydrostatic pressure treatment. Int. J. Food Microbiol. 122, 180-187. Kural, A.G., Kingsley, D.H., Shearer, A.E.H., Chen, H., 2008. Conditions for high pressure inactivation of Vibrio parahaemolyticus in oysters. Int. J. Food Microbiol, 127, 1-5. Liu, T.S., Snoeyenbos, G.H., Carlson, V.L., 1969. Thermal resistance of Salmonella Senftenberg 775W in dry animal feeds. Avian Dis. 13(6), 611-631. Martin, R.E., Cushwa, C.T., 1966. Effects of heat and moisture on leguminous seed. Proc. Annu. Tall Timbers Fire Ecol. Conf. 5, 159-175. Mattick, K.L., Jorgensen, F., Legan, J.D., Lappin-Scott, H., Humphrey, T.J., 2000. Habituation of Salmonella spp. at reduced water activity and its effect on heat tolerance. Appl. Environ. Microbiol. 66, 4921-4925. Meyerowitz, S., 1999. Sprouts: the miracle food. Sproutman Publications, Massachusetts, pp. 57-79. Neetoo, H., Ye, M., Chen, H., 2008. Potential application of high hydrostatic pressure to eliminate Escherichia coli O157:H7 on alfalfa sprouted seeds. Int. J. Food Microbiol. 128, 348-353. Neetoo, H., Chen, H., Pizzolato, T., 2009. Elimination of Escherichia coli O157:H7 from alfalfa seeds through a combination of high hydrostatic pressure and mild heat. Appl. Environ. Microbiol. 75, 1901-1907. Neetoo, H., Chen, H., 2010. Inactivation of Salmonella and Escherichia coli O157:H7 on artificially contaminated alfalfa seeds using high hydrostatic pressure. Food Microbiol. 27, 332-338. Ng, H., Bayne, H.G., Garibaldi, J.A., 1969. Heat resistance of Salmonella: the uniqueness of Salmonella Seftenberg775W. Appl. Microbiol. 17, 78-82. O’ Mahony, M., Cowden, J., Smyth, B., Lynch, D., Hall, M., Rowe, B., Tearle, E.L., Tettmar, R.E., Rampling, A.S., Coles, M., Gilbert, R.J., Kingcott, R.J., Barlett, C.L.R., 1990. An outbreak of Salmonella Saintpaul infection associated with bean sprouts. Epidemiol. Infect. 104, 229-235. Ponce, E., Pla, R., Capellas, M., Guamis, B., Mor-Mur, M., 1998. Inactivation of Escherichia coli inoculated in liquid whole egg by high hydrostatic pressure. Food Microbiol. 15, 265-272.

231
Ponce, E., Pla, R., Sendra, E., Guamis, B., Mor-Mur, M., 1999. Destruction of Salmonella enteritidis inoculated in liquid whole egg by high hydrostatic pressure: comparative study in selective and non-selective media. Food Microbiol. 16, 357-365. Ponka, A., Anderson, Y., Sitonen, A., DeJong, B., Jahkola, M., Haikala, O., Kuhmonen, A., Pakkala, P., 1995. Salmonella in alfalfa sprouts. Lancet. 345, 462–463. Proctor, M.E., Hamacher, M., Tortorello, M.L., Archer, J.R., Davis, J.P., 2001. Multistate outbreak of Salmonella serovar Muenchen infections associated with alfalfa sprouts grown from seeds pretreated with calcium hypochlorite. J. Clin. Microbiol. 39, 3461-3465. Puohiniemi, R., Heiskanen, T., Sitonen, A., 1997. Molecular epidemiology of two international sprout-borne Salmonella outbreaks. J. Clin. Microbiol. 35, 2487–2491. Rajkowski, K.T., Thayer, D.W., 2001. Alfalfa seed germination and yield ratio and alfalfa sprout microbial keeping quality following irradiation of seeds and sprouts. J. Food Prot. 64, 1988-1995. Riemann, H., 1968. Effect of water activity on the heat resistance of Salmonella in ‘dry’ materials. Appl. Microbiol. 16, 1621–1622. Romling, U., Bian, Z., Hammar, M., Sierralta, W.D., Normark, S. 1998. Curli fibers are highly conserved between Salmonella Typhimurium and Escherichia coli with respect to operon structure and regulation. J. Bacteriol. 180, 722-731. Staker, E.V., 1925. The effect of dry heat on alfalfa and its adulterants. J. Am. Soc. Agron. 17, 32-40. Stewart, G., 1926. Effect of color of seed, of scarification, and of dry heat on the germination of alfalfa seed and some of its impurities. J. Am. Soc. Agron. 18, 743-761. Sumner, S.S., Sandros, T.M., Harmon, M.C., Scott, V.N., Bernard, D.T., 1991. Heat resistance of Salmonella Typhimurium and Listeria monocytogenes in sucrose solutions of various water activities. J. Food Sci. 56, 1741-1743. Taormina, P.J., Beuchat, L.R., Slutsker, L., 1999. Infections associated with eating seed sprouts: an international concern. Emerg. Infect. Dis. 5, 626-634. Van Beneden, C.A., Keene, W.E., Stang, R.A., Werker, D.H., King, A.S., Mahon, B., Hedberg, K., Bell, A., Kelly, M.T., Balan, V.K., MacKenzie, W.R., Fleming, D., 1999. Multinational outbreak of Salmonella Enterica serotype Newport infections due to contaminated alfalfa sprouts. J. Am. Med. Assoc. 281, 158-162. Wang, B.S.P., Berjak, P., 2000. Beneficial effects of moist chilling on the seeds of black spruce (Picea mariana [Mill.] B.S.P.). Ann. Bot. 86, 29–36.
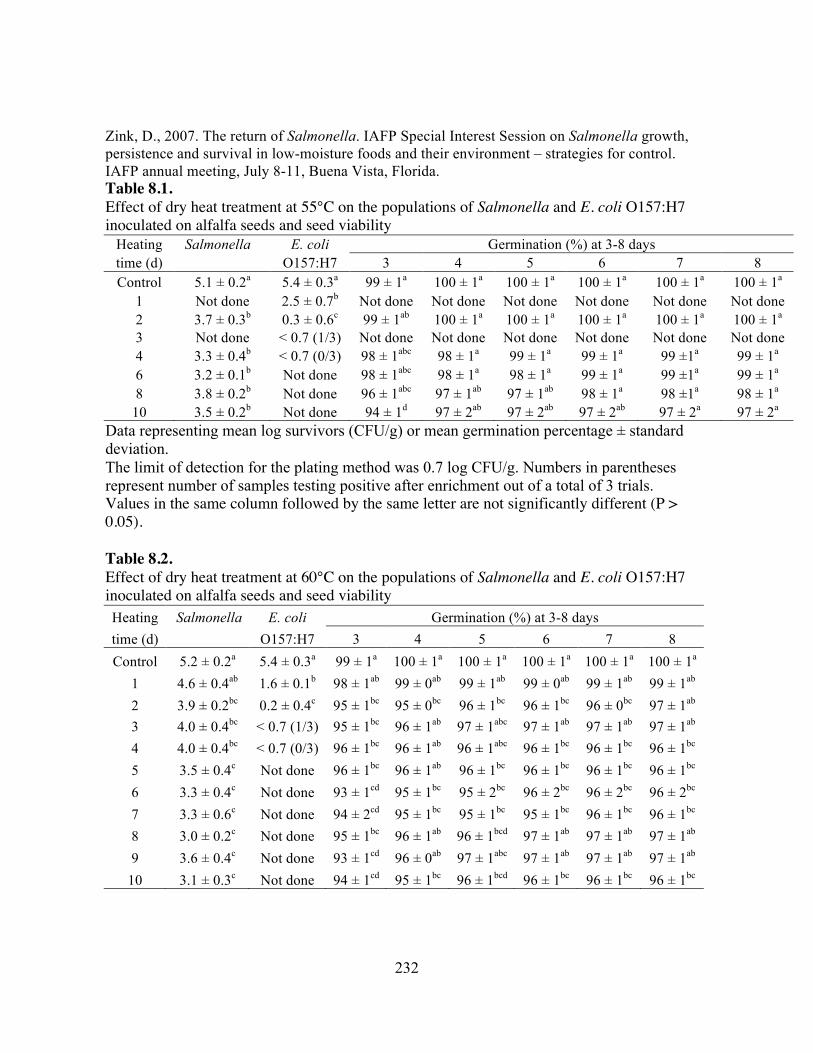
232
Zink, D., 2007. The return of Salmonella. IAFP Special Interest Session on Salmonella growth, persistence and survival in low-moisture foods and their environment – strategies for control. IAFP annual meeting, July 8-11, Buena Vista, Florida. Table 8.1. Effect of dry heat treatment at 55°C on the populations of Salmonella and E. coli O157:H7 inoculated on alfalfa seeds and seed viability
Heating Salmonella E. coli Germination (%) at 3-8 days time (d) O157:H7 3 4 5 6 7 8 Control 5.1 ± 0.2a 5.4 ± 0.3a 99 ± 1a 100 ± 1a 100 ± 1a 100 ± 1a 100 ± 1a 100 ± 1a
1 Not done 2.5 ± 0.7b Not done Not done Not done Not done Not done Not done 2 3.7 ± 0.3b 0.3 ± 0.6c 99 ± 1ab 100 ± 1a 100 ± 1a 100 ± 1a 100 ± 1a 100 ± 1a 3 Not done < 0.7 (1/3) Not done Not done Not done Not done Not done Not done 4 3.3 ± 0.4b < 0.7 (0/3) 98 ± 1abc 98 ± 1a 99 ± 1a 99 ± 1a 99 ±1a 99 ± 1a 6 3.2 ± 0.1b Not done 98 ± 1abc 98 ± 1a 98 ± 1a 99 ± 1a 99 ±1a 99 ± 1a 8 3.8 ± 0.2b Not done 96 ± 1abc 97 ± 1ab 97 ± 1ab 98 ± 1a 98 ±1a 98 ± 1a
10 3.5 ± 0.2b Not done 94 ± 1d 97 ± 2ab 97 ± 2ab 97 ± 2ab 97 ± 2a 97 ± 2a Data representing mean log survivors (CFU/g) or mean germination percentage ± standard deviation. The limit of detection for the plating method was 0.7 log CFU/g. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials. Values in the same column followed by the same letter are not significantly different (P > 0.05). Table 8.2. Effect of dry heat treatment at 60°C on the populations of Salmonella and E. coli O157:H7 inoculated on alfalfa seeds and seed viability Heating Salmonella E. coli Germination (%) at 3-8 days time (d) O157:H7 3 4 5 6 7 8 Control 5.2 ± 0.2a 5.4 ± 0.3a 99 ± 1a 100 ± 1a 100 ± 1a 100 ± 1a 100 ± 1a 100 ± 1a
1 4.6 ± 0.4ab 1.6 ± 0.1b 98 ± 1ab 99 ± 0ab 99 ± 1ab 99 ± 0ab 99 ± 1ab 99 ± 1ab 2 3.9 ± 0.2bc 0.2 ± 0.4c 95 ± 1bc 95 ± 0bc 96 ± 1bc 96 ± 1bc 96 ± 0bc 97 ± 1ab 3 4.0 ± 0.4bc < 0.7 (1/3) 95 ± 1bc 96 ± 1ab 97 ± 1abc 97 ± 1ab 97 ± 1ab 97 ± 1ab 4 4.0 ± 0.4bc < 0.7 (0/3) 96 ± 1bc 96 ± 1ab 96 ± 1abc 96 ± 1bc 96 ± 1bc 96 ± 1bc 5 3.5 ± 0.4c Not done 96 ± 1bc 96 ± 1ab 96 ± 1bc 96 ± 1bc 96 ± 1bc 96 ± 1bc 6 3.3 ± 0.4c Not done 93 ± 1cd 95 ± 1bc 95 ± 2bc 96 ± 2bc 96 ± 2bc 96 ± 2bc 7 3.3 ± 0.6c Not done 94 ± 2cd 95 ± 1bc 95 ± 1bc 95 ± 1bc 96 ± 1bc 96 ± 1bc 8 3.0 ± 0.2c Not done 95 ± 1bc 96 ± 1ab 96 ± 1bcd 97 ± 1ab 97 ± 1ab 97 ± 1ab 9 3.6 ± 0.4c Not done 93 ± 1cd 96 ± 0ab 97 ± 1abc 97 ± 1ab 97 ± 1ab 97 ± 1ab
10 3.1 ± 0.3c Not done 94 ± 1cd 95 ± 1bc 96 ± 1bcd 96 ± 1bc 96 ± 1bc 96 ± 1bc
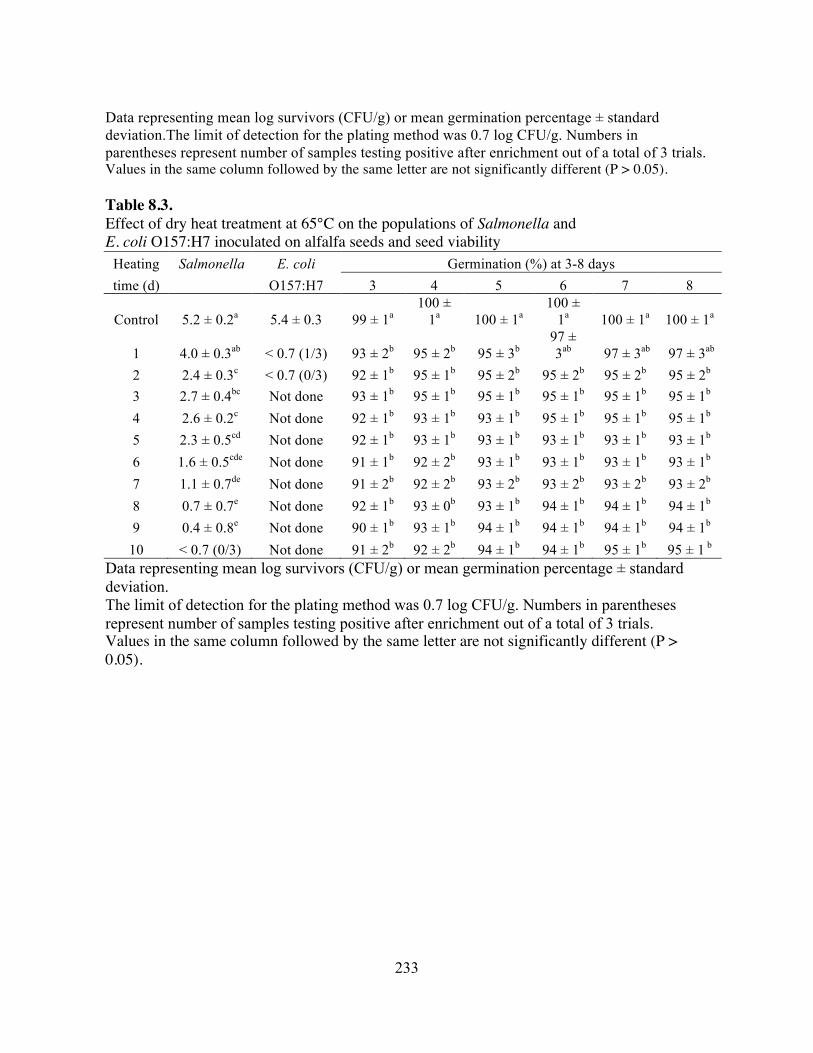
233
Data representing mean log survivors (CFU/g) or mean germination percentage ± standard deviation.The limit of detection for the plating method was 0.7 log CFU/g. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials. Values in the same column followed by the same letter are not significantly different (P > 0.05). Table 8.3. Effect of dry heat treatment at 65°C on the populations of Salmonella and E. coli O157:H7 inoculated on alfalfa seeds and seed viability
Heating Salmonella E. coli Germination (%) at 3-8 days time (d) O157:H7 3 4 5 6 7 8
Control 5.2 ± 0.2a 5.4 ± 0.3 99 ± 1a 100 ±
1a 100 ± 1a 100 ±
1a 100 ± 1a 100 ± 1a
1 4.0 ± 0.3ab < 0.7 (1/3) 93 ± 2b 95 ± 2b 95 ± 3b 97 ± 3ab 97 ± 3ab 97 ± 3ab
2 2.4 ± 0.3c < 0.7 (0/3) 92 ± 1b 95 ± 1b 95 ± 2b 95 ± 2b 95 ± 2b 95 ± 2b 3 2.7 ± 0.4bc Not done 93 ± 1b 95 ± 1b 95 ± 1b 95 ± 1b 95 ± 1b 95 ± 1b 4 2.6 ± 0.2c Not done 92 ± 1b 93 ± 1b 93 ± 1b 95 ± 1b 95 ± 1b 95 ± 1b 5 2.3 ± 0.5cd Not done 92 ± 1b 93 ± 1b 93 ± 1b 93 ± 1b 93 ± 1b 93 ± 1b 6 1.6 ± 0.5cde Not done 91 ± 1b 92 ± 2b 93 ± 1b 93 ± 1b 93 ± 1b 93 ± 1b 7 1.1 ± 0.7de Not done 91 ± 2b 92 ± 2b 93 ± 2b 93 ± 2b 93 ± 2b 93 ± 2b 8 0.7 ± 0.7e Not done 92 ± 1b 93 ± 0b 93 ± 1b 94 ± 1b 94 ± 1b 94 ± 1b 9 0.4 ± 0.8e Not done 90 ± 1b 93 ± 1b 94 ± 1b 94 ± 1b 94 ± 1b 94 ± 1b
10 < 0.7 (0/3) Not done 91 ± 2b 92 ± 2b 94 ± 1b 94 ± 1b 95 ± 1b 95 ± 1 b Data representing mean log survivors (CFU/g) or mean germination percentage ± standard deviation. The limit of detection for the plating method was 0.7 log CFU/g. Numbers in parentheses represent number of samples testing positive after enrichment out of a total of 3 trials. Values in the same column followed by the same letter are not significantly different (P > 0.05).
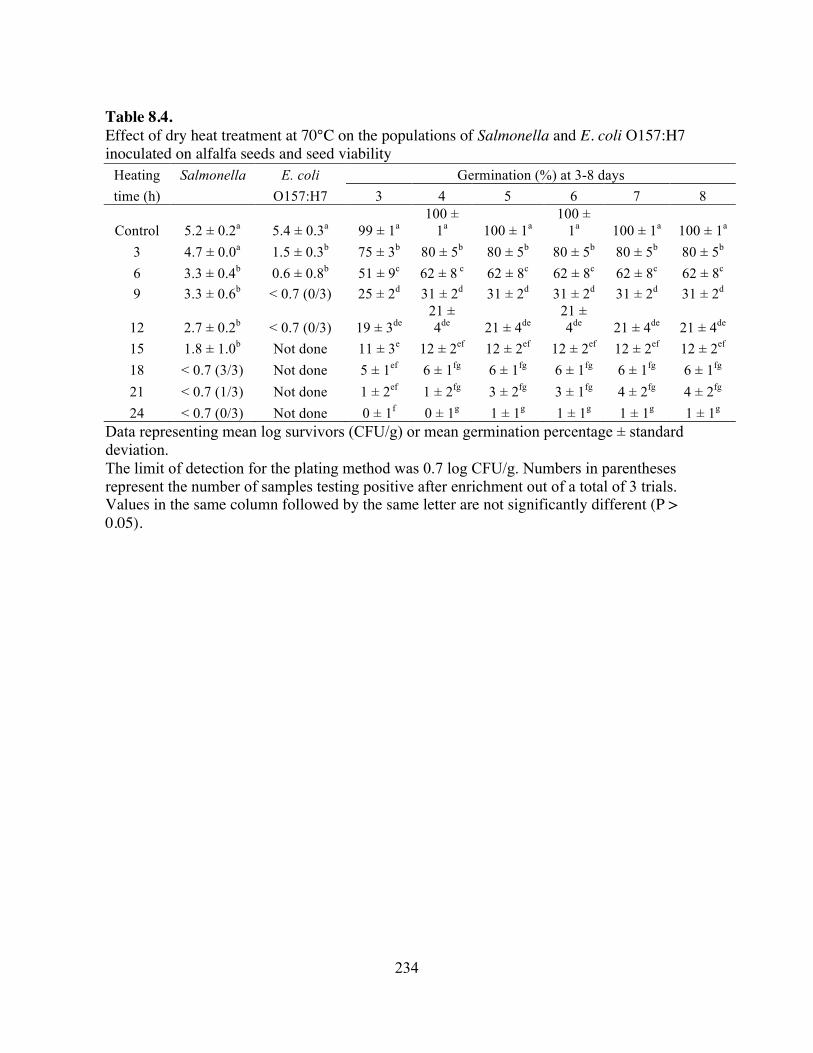
234
Table 8.4. Effect of dry heat treatment at 70°C on the populations of Salmonella and E. coli O157:H7 inoculated on alfalfa seeds and seed viability
Heating Salmonella E. coli Germination (%) at 3-8 days time (h) O157:H7 3 4 5 6 7 8
Control 5.2 ± 0.2a 5.4 ± 0.3a 99 ± 1a 100 ±
1a 100 ± 1a 100 ±
1a 100 ± 1a 100 ± 1a 3 4.7 ± 0.0a 1.5 ± 0.3b 75 ± 3b 80 ± 5b 80 ± 5b 80 ± 5b 80 ± 5b 80 ± 5b 6 3.3 ± 0.4b 0.6 ± 0.8b 51 ± 9c 62 ± 8 c 62 ± 8c 62 ± 8c 62 ± 8c 62 ± 8c 9 3.3 ± 0.6b < 0.7 (0/3) 25 ± 2d 31 ± 2d 31 ± 2d 31 ± 2d 31 ± 2d 31 ± 2d
12 2.7 ± 0.2b < 0.7 (0/3) 19 ± 3de 21 ± 4de 21 ± 4de
21 ± 4de 21 ± 4de 21 ± 4de
15 1.8 ± 1.0b Not done 11 ± 3e 12 ± 2ef 12 ± 2ef 12 ± 2ef 12 ± 2ef 12 ± 2ef 18 < 0.7 (3/3) Not done 5 ± 1ef 6 ± 1fg 6 ± 1fg 6 ± 1fg 6 ± 1fg 6 ± 1fg 21 < 0.7 (1/3) Not done 1 ± 2ef 1 ± 2fg 3 ± 2fg 3 ± 1fg 4 ± 2fg 4 ± 2fg 24 < 0.7 (0/3) Not done 0 ± 1f 0 ± 1g 1 ± 1g 1 ± 1g 1 ± 1g 1 ± 1g
Data representing mean log survivors (CFU/g) or mean germination percentage ± standard deviation. The limit of detection for the plating method was 0.7 log CFU/g. Numbers in parentheses represent the number of samples testing positive after enrichment out of a total of 3 trials. Values in the same column followed by the same letter are not significantly different (P > 0.05).

235
Table 8.5. Effect of combined application of dry heat treatment with pressure treatment on the inactivation Salmonella spp. with an initial population of 5.8 log CFU/g. 600 MPa for 2 min at Heat treatment 20°C 35°C 1 day at 55°C 1.5 ± 1.2 < 0.8 (3/3) 1 day at 60°C < 0.8 (1/3) < 0.8 (0/3) 2 days at 55°C 1.5 ± 1.2 < 0.8 (3/3) 2 days at 60°C < 0.8 (1/3) Not done 3 days at 55°C 1.0 ± 0.3 < 0.8 (1/3) 3 days at 60°C < 0.8 (1/3) Not done 4 days at 55°C < 0.8 (1/3) < 0.8 (0/3) 4 days at 60°C < 0.8 (1/3) Not done
3 h at 65°C 1.6 ± 1.1 1.2 ± 0.7 3 h at 70°C < 0.8 (3/3) < 0.8 (1/3) 6 h at 65°C 0.9 ± 0.6 1.1 ± 0.6 6 h at 70°C < 0.8 (3/3) < 0.8 (0/3) 9 h at 65°C < 0.8 (3/3) < 0.8 (3/3) 9 h at 70°C < 0.8 (1/3) Not done 12 h at 65°C < 0.8 (1/3) < 0.8 (0/3) 12 h at 70°C Not done Not done
Data representing mean log survivors (CFU/g) ± standard deviation. The limit of detection for the plating method was 0.8 log CFU/g. Numbers in parentheses represent the number of samples testing positive after enrichment out of a total of 3 trials.
Related Documents











