Cytokine components and mucosal immunity in the oviduct of Xenopus laevis (amphibia, pipidae) Silke Jantra a , Luana Paulesu a,⇑ , Mario Lo Valvo b , Francesco Lillo b , Francesca Ietta a , Anna Maria Avanzati c , Roberta Romagnoli a , Nicoletta Bechi a , Rossana Brizzi d a Department of Physiology, University of Siena, Via A. Moro, 53100 Siena, Italy b Department of Environmental Biology and Biodiversity, University of Palermo, Via Archirafi 18, 90123 Palermo, Italy c Department of Evolutionary Biology, University of Siena, Via A. Moro, 53100 Siena, Italy d Department of Evolutionary Biology ‘‘Leo Pardi’’, University of Florence, Via Romana 17, 50125 Florence, Italy article info Article history: Received 10 January 2011 Revised 18 July 2011 Accepted 20 July 2011 Available online 27 July 2011 Keywords: Female reproductive tissues Interleukin-1 Interleukin-1 receptor type 1 Macrophage migration inhibitory factor Xenopus laevis abstract Most studies on the mucosal immunity in female reproductive tissues have been performed in mammals. In all species, apart from their reproductive strategies, immunity in the genital mucosa is required to defend the host against luminal pathogens. In this study we investigated the role of the innate immunity of the oviductal mucosa of Xenopus laevis, an amphibian characterized by external fertilization. In particular we examined the expression and local- ization of Interleukin-1b (IL1B), Macrophage migration inhibitory factor (MIF) and Interleukin-1 receptor type 1 (IL1R1) in different oviductal portions including an upper glandular region, an intermediate and a lower aglandular region (the ovisac). Tissues were examined by immunohistochemistry and western blot using polyclonal antibodies against human molecules. IL1B, MIF and IL1R1 were all shown in the three oviductal regions examined, albeit with a general increase towards the external environment. A substantial difference among the cytokine components was also observed mainly in the epithelium of the glandular and intermediate regions. Specifically, all three molecules were expressed by the luminal ciliated cells while only IL1R1 was present in the uncil- iated cells at the bottom of the epithelial ingrowths. The expression of IL1R1 in these cells appeared as a continuous layer separating the epithelium from the underlying tissues. While supporting the role of the innate immune system for host’s defense against pathogens, the pecu- liar distribution of the cytokine components in the oviduct of X. laevis suggests novel immunologic strat- egies useful to assure gland secretion essential for egg formation and fertilization. Ó 2011 Elsevier Inc. All rights reserved. 1. Introduction The mucosa of the female reproductive tract as well as the one of the intestinal and bronchial tracts interfaces with a lumen exposed to an extensive variety of both commensal and pathogenic microbes coming from the external environment [4]. To control the bacterial antigenic load, the mucosal epithelium of the female reproductive tract, like that of other mucosal sites, develops defense mechanisms mainly based on expression of receptors specific for pathogen associated molecular patterns, e.g., Toll-like receptors (TLRs), [4,33]. TLRs, named after the Drosophila Toll gene, are considered major mediators of the innate immune response from insects to mammals [12,15,23,26,31, 34,51]. When microbial antigens come in contact with TLRs, cells of the mucosal epithelium are activated to protect the host as well as to alert other cells of the underlying tissues through secretion of cyto- kines [1,14,52]. Cytokines are soluble mediators of the innate immune re- sponse, an ancient immunologic defense mechanism present throughout vertebrates up to mammals [50]. The interleukin-1 (IL1) system is a widely studied cytokine family with pleiotropic functions in a variety of cell types (reviewed in [10]). It consists of different components mainly including two bioactive ligands (IL1A and IL1B) and two receptors: IL1 receptor type I (IL1R1) and type II (IL1R2) [13,32,47]. Only the binding to IL1R1 results in signal transduction [46] whereas IL1R2 acts as a competitor [9,11]. 0016-6480/$ - see front matter Ó 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2011.07.003 ⇑ Corresponding author. E-mail addresses: [email protected] (S. Jantra), [email protected] (L. Paulesu), [email protected] (M. Lo Valvo), [email protected] (F. Lillo), [email protected] (F. Ietta), [email protected] (A.M. Avanzati), [email protected] (R. Romagnoli), [email protected] (N. Bechi), rossana.brizzi@unifi.it (R. Brizzi). General and Comparative Endocrinology 173 (2011) 454–460 Contents lists available at ScienceDirect General and Comparative Endocrinology journal homepage: www.elsevier.com/locate/ygcen

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

General and Comparative Endocrinology 173 (2011) 454–460
Contents lists available at ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Cytokine components and mucosal immunity in the oviduct of Xenopus laevis(amphibia, pipidae)
Silke Jantra a, Luana Paulesu a,⇑, Mario Lo Valvo b, Francesco Lillo b, Francesca Ietta a,Anna Maria Avanzati c, Roberta Romagnoli a, Nicoletta Bechi a, Rossana Brizzi d
a Department of Physiology, University of Siena, Via A. Moro, 53100 Siena, Italyb Department of Environmental Biology and Biodiversity, University of Palermo, Via Archirafi 18, 90123 Palermo, Italyc Department of Evolutionary Biology, University of Siena, Via A. Moro, 53100 Siena, Italyd Department of Evolutionary Biology ‘‘Leo Pardi’’, University of Florence, Via Romana 17, 50125 Florence, Italy
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 January 2011Revised 18 July 2011Accepted 20 July 2011Available online 27 July 2011
Keywords:Female reproductive tissuesInterleukin-1Interleukin-1 receptor type 1Macrophage migration inhibitory factorXenopus laevis
0016-6480/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.ygcen.2011.07.003
⇑ Corresponding author.E-mail addresses: [email protected] (S. Jantra), p
[email protected] (M. Lo Valvo), francesco.lillo@gmai(F. Ietta), [email protected] (A.M. Avanzati), [email protected] (N. Bechi), [email protected] (R. B
Most studies on the mucosal immunity in female reproductive tissues have been performed in mammals.In all species, apart from their reproductive strategies, immunity in the genital mucosa is required todefend the host against luminal pathogens.
In this study we investigated the role of the innate immunity of the oviductal mucosa of Xenopus laevis,an amphibian characterized by external fertilization. In particular we examined the expression and local-ization of Interleukin-1b (IL1B), Macrophage migration inhibitory factor (MIF) and Interleukin-1 receptortype 1 (IL1R1) in different oviductal portions including an upper glandular region, an intermediate and alower aglandular region (the ovisac). Tissues were examined by immunohistochemistry and western blotusing polyclonal antibodies against human molecules.
IL1B, MIF and IL1R1 were all shown in the three oviductal regions examined, albeit with a generalincrease towards the external environment. A substantial difference among the cytokine componentswas also observed mainly in the epithelium of the glandular and intermediate regions. Specifically, allthree molecules were expressed by the luminal ciliated cells while only IL1R1 was present in the uncil-iated cells at the bottom of the epithelial ingrowths. The expression of IL1R1 in these cells appeared as acontinuous layer separating the epithelium from the underlying tissues.
While supporting the role of the innate immune system for host’s defense against pathogens, the pecu-liar distribution of the cytokine components in the oviduct of X. laevis suggests novel immunologic strat-egies useful to assure gland secretion essential for egg formation and fertilization.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
The mucosa of the female reproductive tract as well as the oneof the intestinal and bronchial tracts interfaces with a lumenexposed to an extensive variety of both commensal and pathogenicmicrobes coming from the external environment [4].
To control the bacterial antigenic load, the mucosal epitheliumof the female reproductive tract, like that of other mucosal sites,develops defense mechanisms mainly based on expression ofreceptors specific for pathogen associated molecular patterns,e.g., Toll-like receptors (TLRs), [4,33]. TLRs, named after the
ll rights reserved.
[email protected] (L. Paulesu),l.com (F. Lillo), [email protected]@unisi.it (R. Romagnoli),rizzi).
Drosophila Toll gene, are considered major mediators of the innateimmune response from insects to mammals [12,15,23,26,31,34,51].
When microbial antigens come in contact with TLRs, cells of themucosal epithelium are activated to protect the host as well as toalert other cells of the underlying tissues through secretion of cyto-kines [1,14,52].
Cytokines are soluble mediators of the innate immune re-sponse, an ancient immunologic defense mechanism presentthroughout vertebrates up to mammals [50]. The interleukin-1(IL1) system is a widely studied cytokine family with pleiotropicfunctions in a variety of cell types (reviewed in [10]). It consistsof different components mainly including two bioactive ligands(IL1A and IL1B) and two receptors: IL1 receptor type I (IL1R1)and type II (IL1R2) [13,32,47]. Only the binding to IL1R1 resultsin signal transduction [46] whereas IL1R2 acts as a competitor[9,11].

S. Jantra et al. / General and Comparative Endocrinology 173 (2011) 454–460 455
Cloning of IL1B gene has been performed in mammals, birds,amphibians [54] and fishes (reviewed in [5]). The intracytoplasmicregion of IL1R1 resembles that of the corresponding region of TLRscommon to all vertebrates [44]. This region, referred as Toll/IL1R(TIR), has been identified in the TLRs of mammals and other verte-brates (fishes, amphibians and birds) [12,44]. Therefore, binding toIL1R1 produces signals that are similar to those of TLRs acting aseffectors of the innate immune response [44]. The IL1 system,mainly IL1B and IL1R1, has been detected in the female reproduc-tive tissues of different animal groups from mammals to elasmo-branchs [8,37–39,41]. The presence of IL1 system has beenusually associated to maternal immunotolerance towards thesemi-allogeneic embryo (reviewed in [40]). However, in a recentstudy, we showed expression of IL1B and IL1R1 even in the femalereproductive tissues of ovuliparous amphibian species, namelythose with external fertilization [27]. This finding suggested thatthe presence of IL1 system in the female genital tract is primarilyrelated to the defense against microorganisms, regardless of thespecific reproductive mode.
A further cytokine with very well known activities in the innateimmune response is the Macrophage Migration Inhibitory Factor(MIF). The MIF protein is remarkably well conserved throughoutevolution, and its homologs are encoded in different vertebrates,invertebrates and plants (reviewed in [18]). MIF is thought to playa central role in the host response to microbial infections and toregulate the innate immunity through modulation of TLR 4 [45,46].
In mammals, MIF is highly expressed by cells and tissues in di-rect contact with the host’s external environment, including thelungs, the skin, and the epithelial lining of the gastrointestinaland genitourinary tracts [2,3,30]. Several studies showed thatMIF is largely involved in reproduction and its expression in the fe-male reproductive tissues has been mainly related to the establish-ment and maintenance of pregnancy [17,24,37]. Nonetheless, MIFhas never been inquired in the genital apparatus of ovuliparousspecies.
In this paper we examined the expression of the cytokines MIFand IL1B and of the IL1R1 in different portions of the oviduct ofXenopus laevis, a completely aquatic anuran characterized by exter-nal fertilization. The main goal of this study was to investigate therole of the oviductal mucosa in the innate immunity, specifically inrelation to its communication with the external aquatic environ-ment and the necessity to perform an active defense against lumi-nal pathogenic microorganisms.
2. Materials and methods
2.1. Collection of samples and tissue processing
X. laevis specimens were collected during the reproductive sea-son in agriculture ponds near Palermo, Italy, where this species hasfounded a numerous alien population [16]. Investigation on wildspecimens allowed us to exclude possible influences related tocaptivity conditions.
Immediately after their transport to the laboratory, the animalswere sacrificed in overdose of clorobutanol 0.2% for at least 20 min,opened by a midventral incision and the oviducts were isolated.Eight adult females of X. laevis were selected on the basis of thepresence of mature eggs in the ovaries. For this study we examinedthe caudal portion of each oviduct, a segment about 3 cm long,including the last portion of the pars convoluta and the followingovisac (region PC4 and uterus, respectively, described in thisspecies by Yoshizaki [53]. These portions of the right oviduct werefixed in 10% buffered neutral formalin for 24 h, washed inwater and dehydrated in ethanol, and then embedded in paraffinwax for current histologic (Mayer’s hematoxylin–eosin) or
immunohistochemical staining. The corresponding portions ofthe left oviduct were immediately snap-frozen in liquid nitrogenand stored at �80 �C for Western blot analysis to determine themolecular weight of the proteins recognized. The spleen was alsoisolated, snap-frozen and used as a positive control as it is a tissueexpected to express the cytokines analyzed in this study [5,48]. Theexperiments and animal captures were performed with the ap-proval of the institutional committees and the Italian Ministry ofthe Environment (DPN/2D/2006/16274).
2.2. Immunohistochemistry
Formalin-fixed, paraffin embedded, 5 lm sections were depa-raffinated and rehydrated in serial dilutions of ethanol. Antigen re-trieval was carried out by incubating sections in sodium citratebuffer (10 mM, pH 6.0) in a microwave oven at 750 W three timesfor 5 min. Sections were washed in Tris buffer saline (TBS; 50 mMTris–HCl, 150 mM NaCl; [pH 7.6]) and pre-incubated for 20 minwith normal rabbit serum (DAKO, Copenhagen, Denmark) diluted1:10 in TBS to prevent non-specific binding. After incubation over-night at 4 �C with anti-human IL1B (50 lg/ml) (R&D Systems,Abingdon, UK), anti-human IL1R1 (50 lg/ml) (R&D Systems) oranti-human MIF (25 lg/ml) (R&D Systems) goat polyclonal anti-bodies, the slides were rinsed three times in TBS for 5 min andincubated for 30 min with rabbit anti-goat secondary antibody la-beled with biotin at a dilution of 1:500 (DAKO). After being washedanother three times for 5 min in TBS, sections were incubated for30 min with Streptavidin complex/AP (DAKO) diluted 1:300. Thealkaline phosphatase reaction was revealed using naphtol andnew fuchsin as substrate. The endogenous alkaline phosphatasewas blocked by adding 1 mM levamisole to the substrate solution.Sections were not counterstained and immediately washed for5 min in running tap water and mounted with aqueous mountingmedium (Aquatex, Merck, Darmstadt, Germany). Negative controlswere performed for each tissue by substituting the primary anti-body with TBS.
Based on previous experience in other non-human species[27,38,39], the specificity of the polyclonal antibodies against hu-man IL1B and MIF was evaluated by pre-adsorption (overnight at4 �C) of the primary antibodies with the human recombinant pro-teins IL1B (Abcam, Cambridge, UK) and MIF (R&D Systems) at amolar ratio of 1:10.
The specificity of the anti-IL1R1 antibody was confirmed by pre-adsorption on BeWo cells expressing the receptor [25]. BeWo cells(Istituto Zooprofilattico Sperimentale, Brescia, Italy) were culturedin Ham’s F-10 without phenol red (Sigma Chemical Co.) supple-mented with 10% FBS (Biochrom, Berlin, Germany), 100 U/mlpenicillin/streptomycin and 2 mM glutamine (Sigma ChemicalCo.) in 96 well cell culture plates (Becton Dickinson Labware,Franklin Lakes, NJ, USA) in a humidified atmosphere of 20% airand 5% CO2 at 37 �C until 70–80% confluence. Cultures werewashed three times in phosphate buffer saline (PBS) then, incu-bated with 50 lg/ml of the anti-IL1R1 for 1 h at 0 �C. After 5 mincentrifugation at 16,000g at 4 �C, the supernatant was used forimmunohistochemistry.
2.3. Protein extraction
The frozen caudal portion of the left oviduct and spleen tissuesof X. laevis were lysed twice for 60 s in extraction buffer containing5% beta-mercaptoethanol, 5% SDS, 62 mM Tris, pH 6.8. and 0.01%protease inhibitor cocktail (Sigma–Aldrich, St. Louis, MO, USA)using a TURAX homogenizer. After centrifugation at 16,000g for15 min at 4 �C, the supernatant was assayed for total proteincontent (Bradford, Bio-Rad, Richmond, CA, USA) and stored at�80 �C until Western blot analysis for IL1B and MIF.

456 S. Jantra et al. / General and Comparative Endocrinology 173 (2011) 454–460
2.4. Gel electrophoresis and western blotting
Appropriate volumes of protein extracts, adjusted to representthe same amount of total protein (100 lg/lane for IL1B and50 lg/lane for MIF), were separated under reducing condition atroom temperature using the NuPage Novex kit (Invitrogen, Carls-bad, CA, USA) on polyacrylamide gel pre-casting 12% (Invitrogen).After electrophoretic transfer to a PVDF membrane (Hybond-P;Amersham, Biosciences, Little Chalfont, UK) at 100 V for 1 h at4 �C, the membranes were incubated in blocking solution [BS; 5%(w/v) fat-free dry milk in 10 mM PBS, 0.15 M NaCl pH 7.4, and0.1% (v/v) Tween 20] for 1.5 h at room temperature. Anti-humanIL1B (R&D Systems) or anti-human MIF (R&D Systems) goatpolyclonal antibodies diluted in BS (1 lg/ml) were placed on themembrane and incubated at 4 �C overnight in agitation. After threewashings with 0.1% Tween 20 in phosphate buffer saline (PBS) for10 min each, the secondary antibody, conjugated with horseradishperoxidase (HRP) (Bio-Rad), at a dilution of 1:1000, was appliedand the signals on the membranes were detected by West Picochemiluminescent substrate (Pierce, Rockford, USA) according tothe manufacturer’s instructions.
3. Results
3.1. Structure of the oviduct
In the females of most anurans, including X. laevis, the oviductscan be divided into different segments based upon their gross
Fig. 1. Schematic representation of the oviduct of X. laevis and cross-sections of the regioncollects the ovulated eggs from the coelomic cavity, a pars convolute, which represenoviposition. The shaded areas are detailed in B and C. (B) The different oviductal regionintermediate region (IR), characterized by strong gland reduction and the aglandular ovis(hematoxylin–eosin staining). (C I and II): glandular region showing pluristratified, tubshows ridges covered by luminal ciliated cells. Narrow ingrowths (asterisks) alternate topoint to gland lumina. (C III and IV): Intermediate region characterized by strong glaningrowths (asterisk) consists of unciliated epithelial cells (arrowheads in C IV). (C V and Vshape, where ciliated and unciliated cells form a single layer. Arrows point to blood capil
morphology (Fig. 1A). The uppermost segment, thin and straight,opens into the coelomic cavity and is defined pars recta. The fol-lowing segment, notably longer and repeatedly folded, pars convo-luta, is characterized by the presence of numerous tubular glandslying deep in the connective tissue between the epithelium andthe serosa. These glands supply jelly envelopes to the eggs duringtheir passage through the oviduct. The third segment, termed ovi-sac or uterus, is an aglandular region that opens directly into thecloaca. We specifically studied the lowest tract of the pars convolu-ta and the ovisac. Between these two regions we arbitrarily distin-guished an intermediate region, characterized by a strongreduction in the gland number. Therefore, the oviductal portionsstudied included: a glandular region (GR), an intermediate region(IR) and the ovisac, (Fig. 1B). These different regions were also rec-ognizable by their characteristics in mucosal morphology (Fig. 1CI–VI).
The epithelium of the glandular region showed low ridgesintercalated by narrow ingrowths (Fig. 1C I and II). The ridgeswere covered by ciliated cells, whereas unciliated cells weredistributed at the bottom of the epithelial ingrowths (Fig. 1C II).Just at the bottom of these ingrowths opened the glands, eachcomposed of cells filled with a granular product and arrangedaround a narrow lumen (Fig. 1C I and II). A rich capillary networkramified from the peripheral muscular sheet of the oviduct to themucosal folds.
In the intermediate region, the residual gland layers werecoated by higher ciliated folds, reciprocally separated by saccularingrowths (Fig. 1C III and IV) lined by unciliated cells (Fig. 1C IV).
s examined. (A) The whole oviduct includes three main portions: a pars recta, whichts the main body of the oviduct, and the ovisac, where eggs accumulate before
s examined: a glandular region (GR) from the lowest tract of the pars convoluta, anac. (C) Cross-sections of the regions examined and observed under light microscopeular glands, whose cells are filled with granular product. The oviductal epitheliumthe ridges and their bottom is lined by unciliated cells (arrowheads in C II). Arrows
d reduction and higher ciliated folds along the luminal surface. The bottom of theI): The ovisac lacks glands and its epithelium is arranged in folds of various size andlaries. ct = connective tissue, gl = glands, lu = oviductal lumen, mu = muscular sheet.

S. Jantra et al. / General and Comparative Endocrinology 173 (2011) 454–460 457
The ovisac completely lacked glands and its epithelium was ar-ranged in folds of various height and appearance, where ciliatedand unciliated cells alternated in a single layer (Fig. 1C V and VI).The connective and muscle tissue layers were thicker than in theupper oviductal regions and included numerous and large bloodvessels (Fig. 1C V and VI).
3.2. Immunoreactivity for IL1B, IL1R1 and MIF
Immunoreactivity for IL1B, IL1R1 and MIF was observed, albeitwith a variable pattern, in sections of the three oviductal regionsstudied. Results are reported in Table 1 and Fig. 2.
In the glandular and in the intermediate region, the epitheliumshowed expression for IL1B (Fig. 2A and D), IL1R1 (Fig. 2B and E)and MIF (Fig. 2C and F) although with different localization. Specif-ically, the luminal ciliated cells showed a distinct immunoreactiv-ity for IL1B and MIF mostly in their apical portion (Fig. 2A and C),whereas these cells were weakly positive for IL1R1 (Fig. 2B). TheIL1R1 was localized in particular in the bottom unciliated cells atthe base of the epithelial ridges and ingrowths (Fig. 2B and E).An increasing immunoreactivity for the three cytokine compo-nents was observed in the epithelium from the glandular to theintermediate region. In both these regions, the glandular ducts aswell as the connective and muscle tissues revealed immunoreac-tivity mainly for IL1R1 and MIF (Fig. 2B, C and F and Table 1),whereas the endothelium was only scarcely immunoreactive andno staining was observed for any of the proteins examined in theglandular cells (Table 1).
In the ovisac, immunoreactivity for the three molecules waswidespread throughout the cytoplasm of most of the epithelialcells, not distinguishable in luminal and bottom cell types in thisgenital tract. Also in the endothelium, connective tissue and mus-cle layers immunostaining was stronger than in the two upper ovi-ductal regions (Fig. 2G–I).
Specificity of the antibodies used and negative controls areshown in Fig. 2J–L. A substantial decrease in tissue immunostain-ing was observed after pre-absorption of the anti-human IL1Band MIF antibodies with the specific recombinant proteins(Fig. 2J–L). Similarly, immunostaining for IL1R1 was drastically re-duced by pre-adsorption of the anti-human IL1R1 antibody onBeWo cells in culture (Fig. 2K). No staining at all was observedsubstituting the primary antibodies with TBS (negative controls)(see inserts in Fig. 2J–L).
3.3. Western blot analysis
Western blot analysis using polyclonal antibodies againsthuman IL1B and MIF was performed in tissue lysates to identifythe molecular weight of the immunoreactive molecules in X. laevis.The specificity of the anti-human IL1R1antibody in X. laevis wasalready reported by Jelaso et al. [28] and a band of 80.6 kDacorresponding to the membrane-bound putative frog protein wasrevealed.
Table 1Cytokine components in different regions of the oviduct in X. laevis.
Epithelium Glandular
IL1B IL1R1 MIF IL1B
LC BU LC BU LC BU
Glandular region + � +/� ++ + � �Intermediate region ++ � + +++ ++ � +/–Ovisaca +++ +++ +++ – –
Sections of the three different oviductal regions were examined by light microscopy and i(�); weak staining (+/�); moderate staining (+); medium staining (++); intense staining (
a In the ovisac epithelial cells are not distinguishable in luminal and bottom ones.
As shown in Fig. 3A, the anti-human IL1B revealed a band ofapproximately 17 kDa in the oviduct and spleen of X. laevis, corre-sponding to the molecular weight of the human recombinant pro-tein, hrIL1B. The anti-human MIF showed a band of 12.5 kDa inoviductal and spleen lysates of X. laevis, corresponding to themonomeric recombinant human MIF protein, rhMIF (Fig. 3B). MIFimmunoreactivity at approximately 25 and 36 kDa are consistentwith the dimeric and trimeric form, respectively.
4. Discussion
As in the intestinal and bronchial tracts, epithelial cells of thefemale reproductive tissues form a non-specific immunologic bar-rier for host protection against pathogens [6,20,21,36,43].
While much attention has been paid on innate immunity in thelungs [21,36] and intestine [6,7,20], fewer studies have investi-gated the female reproductive tract. These studies, mainly per-formed on mammals, demonstrated that the epithelial cells ofthis apparatus are exposed to a bacterial charge not previously val-ued [52]. Furthermore, the presence in the female genital tract offunctional TLRs, has suggested the capability of microbial detection[22]. These receptors are differentially expressed in distinct com-partments of the human reproductive apparatus i.e. fallopiantubes, uterine endometrium, cervix and ectocervix, reflecting dif-ferences in microbial exposure [42].
All these studies clearly demonstrate the importance of themucosal immune system in the female reproductive tract. How-ever, they do not explain how this immunity can provide defenseagainst pathogens without rejecting the embryo, an immunologicdistinct organism. In fact, in mammals as well as in all other vivip-arous species, the innate immune system of the maternal genitalapparatus is required to tolerate paternal-derived antigens presenton sperm, fertilized eggs and embryos [40]. On the other hand, inthe ovuliparous species, in which the eggs are released from the fe-male reproductive tract and their fertilization and embryonicdevelopment take place in the external environment there is nocontact between embryonic antigens and maternal tissues. Thesespecies can therefore be considered a natural negative control instudies on maternal–fetal immunotolerance [27,40] and a suitablemodel to investigate the role of the oviductal mucosa in thedefense against pathogens.
For this purpose, we performed studies on the oviduct of theaquatic amphibian X. laevis, whose reproduction is based onexternal fertilization. In particular, we analyzed expression of thecytokines IL1B and MIF and of the IL1 receptor in distinct, longitu-dinal portions of the oviduct, progressively closer to the externalenvironment. These included an upper glandular region, an inter-mediate and a lower aglandular region, the ovisac that directlyopens into the cloaca.
Our findings showed that all three cytokine components werepresent in the different oviductal regions examined albeit with ageneral increase toward the external environment. Immunoreac-
duct Endothelium Connective/muscle tissue
IL1R1 MIF IL1B IL1R1 MIF IL1B IL1R1 MIF
+/� + +/� � +/� � +/� +/�+ ++ +/� � � + + +/�– – – – +/� + + ++ ++ ++
mmunoreactivity was estimated on a semi-quantitative ranking system: no staining+++); LC: luminal ciliated cells; BU: bottom unciliated cells. (– –): Absence of glands.
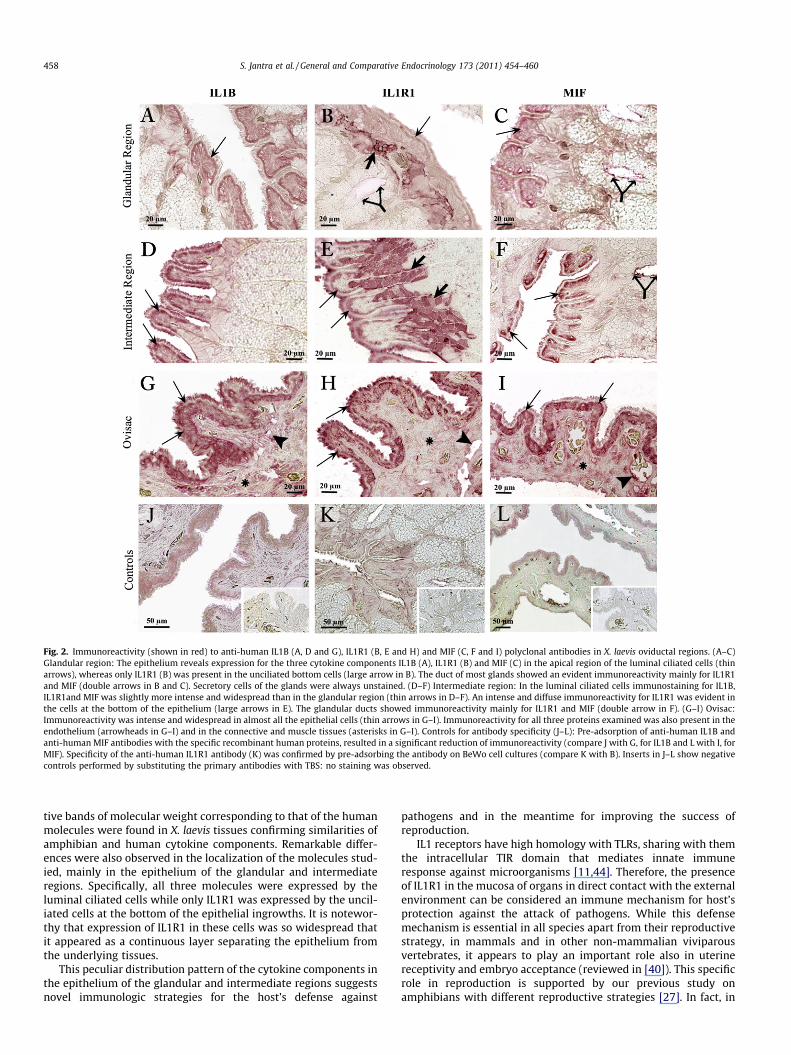
Fig. 2. Immunoreactivity (shown in red) to anti-human IL1B (A, D and G), IL1R1 (B, E and H) and MIF (C, F and I) polyclonal antibodies in X. laevis oviductal regions. (A–C)Glandular region: The epithelium reveals expression for the three cytokine components IL1B (A), IL1R1 (B) and MIF (C) in the apical region of the luminal ciliated cells (thinarrows), whereas only IL1R1 (B) was present in the unciliated bottom cells (large arrow in B). The duct of most glands showed an evident immunoreactivity mainly for IL1R1and MIF (double arrows in B and C). Secretory cells of the glands were always unstained. (D–F) Intermediate region: In the luminal ciliated cells immunostaining for IL1B,IL1R1and MIF was slightly more intense and widespread than in the glandular region (thin arrows in D–F). An intense and diffuse immunoreactivity for IL1R1 was evident inthe cells at the bottom of the epithelium (large arrows in E). The glandular ducts showed immunoreactivity mainly for IL1R1 and MIF (double arrow in F). (G–I) Ovisac:Immunoreactivity was intense and widespread in almost all the epithelial cells (thin arrows in G–I). Immunoreactivity for all three proteins examined was also present in theendothelium (arrowheads in G–I) and in the connective and muscle tissues (asterisks in G–I). Controls for antibody specificity (J–L): Pre-adsorption of anti-human IL1B andanti-human MIF antibodies with the specific recombinant human proteins, resulted in a significant reduction of immunoreactivity (compare J with G, for IL1B and L with I, forMIF). Specificity of the anti-human IL1R1 antibody (K) was confirmed by pre-adsorbing the antibody on BeWo cell cultures (compare K with B). Inserts in J–L show negativecontrols performed by substituting the primary antibodies with TBS: no staining was observed.
458 S. Jantra et al. / General and Comparative Endocrinology 173 (2011) 454–460
tive bands of molecular weight corresponding to that of the humanmolecules were found in X. laevis tissues confirming similarities ofamphibian and human cytokine components. Remarkable differ-ences were also observed in the localization of the molecules stud-ied, mainly in the epithelium of the glandular and intermediateregions. Specifically, all three molecules were expressed by theluminal ciliated cells while only IL1R1 was expressed by the uncil-iated cells at the bottom of the epithelial ingrowths. It is notewor-thy that expression of IL1R1 in these cells was so widespread thatit appeared as a continuous layer separating the epithelium fromthe underlying tissues.
This peculiar distribution pattern of the cytokine components inthe epithelium of the glandular and intermediate regions suggestsnovel immunologic strategies for the host’s defense against
pathogens and in the meantime for improving the success ofreproduction.
IL1 receptors have high homology with TLRs, sharing with themthe intracellular TIR domain that mediates innate immuneresponse against microorganisms [11,44]. Therefore, the presenceof IL1R1 in the mucosa of organs in direct contact with the externalenvironment can be considered an immune mechanism for host’sprotection against the attack of pathogens. While this defensemechanism is essential in all species apart from their reproductivestrategy, in mammals and in other non-mammalian viviparousvertebrates, it appears to play an important role also in uterinereceptivity and embryo acceptance (reviewed in [40]). This specificrole in reproduction is supported by our previous study onamphibians with different reproductive strategies [27]. In fact, in

Fig. 3. Western blot profiles to anti-human IL1B (A) and MIF (B) in oviductal tissuesand spleen of X. laevis. 100 lg/lane for IL1B and 50 lg/lane for MIF of total proteinlysates were run in parallel with human recombinant (rh) IL1B (2 ng) or rhMIF(5 ng). Human placenta (30 lg) was used as positive control for MIF. (A) The anti-human IL1B recognized a band with a molecular weight of approximately 17 kDa inX. laevis tissues, corresponding to that of rhIL1B. (B) Anti-human MIF showed a bandof about 12.5 kDa, corresponding to the predicted monomeric rhMIF protein. Thetwo bands of approximately 25 and 36 kDa are consistent with the dimeric andtrimeric form of MIF, respectively. The position of the molecular weight markers areindicated. The secondary antibody was a rabbit anti-goat conjugated with horse-radish peroxidase (HRP) and the reaction was revealed by a chemiluminescentsubstrate.
S. Jantra et al. / General and Comparative Endocrinology 173 (2011) 454–460 459
the female genital tract of these animals we observed that IL1 sys-tem, namely IL1B and IL1R1, was increasing from ovuliparity tooviparity and viviparity. These findings suggested that the pres-ence of IL1 system in the female reproductive tissues was primarilyrelated to the defense against infections. Its enhancement in ovip-arous and, even more, in viviparous species reflected the need formaternal immunotolerance versus paternal-derived antigens[27,40].
The present study on the ovuliparous X. laevis supports the roleof these immune components in the defense mechanism. The find-ings in different oviductal regions of X. laevis, opened also newhypotheses suggesting a role for IL1 system in reproduction of ovu-liparous species. In particular, the peculiar expression of IL1R1 inthe regions characterized by glands, suggested us that IL1R1 pro-vided an immune barrier at protection of glands. Its distributionappeared in fact as a continuous layer at the bottom of the epithe-lial ingrowths, just near the opening of the oviductal glands. Theseglands supply jelly envelopes to the eggs during their passagethrough the oviduct, therefore playing a fundamental role for eggformation and fertilization in the external environment.
Besides acting as a receptor against pathogens, IL1R1 is essen-tial for IL1 response [10]. Its expression in different cells of the ovi-ductal wall e.g. the luminal epithelium, glandular duct, connective/
muscular tissue and the endothelium, indicates cells targeting theaction of IL1.
As reported in the human uterus by Wira et al. [52], epithelialcells respond to microbial antigens by the release of cytokineswhich then, act as signals on other cell types of the underlying tis-sues. This molecular pathway has been indicated as a second lineof defense against the invading pathogens. Given the wide expres-sion of IL1R1 in different cell types of the oviductal mucosa ofX. laevis, a similar immune mechanism can also be hypothesizedin ovuliparous species.
Comparing the expression of IL1 system in the oviduct ofX. laevis with findings in Bufo bufo [27], it resulted that in this latterspecies immunoreactivity for IL1B and IL1R1 was less widespreadand limited to the apical portion of the luminal epithelium. Thesedifferences in immunoreactivity between the two ovuliparousspecies could reflect their biologic and ecological differences. Infact, B. bufo is almost completely terrestrial, except for a shortaquatic reproductive phase, while X. laevis is entirely aquatic and,as suggested by Todd [49], much more exposed to pathogenicorganisms. The need of a greater presence of mediators of theinnate immune response in X. laevis in relation to a higher patho-genic challenge is supported by the more widespread expressionof the cytokine components in the ovisac. Further studies on theovuliparous anurans characterized by intermediate aquaticity,e.g. some Rana species would be helpful to clarify this point.
Furthermore, investigation on different stages of the reproduc-tive cycle would give important insights on the possible changes incytokine expression and distribution. Preliminary evidence in ourlaboratory indicates that IL1R1 as well as IL1B and MIF are ex-pressed in the oviduct of both, hormone (human chorionic gonado-tropin) treated and untreated X. laevis specimens, corresponding toanimals in the reproductive and non-reproductive phase, respec-tively (data not shown). These data need to be supported by obser-vations on wild specimens.
We did not evaluate MIF receptors because of evidence that,although a high-affinity binding protein, CD74, has been identifiedas cell membrane MIF receptor [35], MIF can also enter the cells byendocytosis [29]. These two different modes of action appear to in-volve different biologic responses with the receptor binding actioninducing growth regulation, apoptosis, and cell cycle control ratherthan inflammatory signal transduction [19].
The evidence of MIF, here shown for the first time in the femalereproductive tract of an ovuliparous species supports the role ofthis cytokine in the innate immunity of tissues contiguous to theexternal environment [45,46].
On the whole, the present findings in the ovuliparous species,X. laevis, appear consistent with two general statements. First,the lower female genital mucosa is capable of activating immuno-logic responses by the secretion and possible action of cytokines.Second, a strategic distribution of IL1R1 near the opening of theoviductal glands represents an immunologic mechanism assuringgland safety and therefore the oviductal secretory activity, essen-tial for egg fertilization and protection outside the body.
References
[1] S. Akira, K. Takeda, Toll-like receptor signaling, Nat. Rev. Immunol. 4 (2004)499–511.
[2] M. Bacher, A. Meinhardt, H.Y. Lan, W. Mu, C.N. Metz, J.A. Chesney, T. Calandra,D. Gemsa, T. Donnelly, R.C. Atkins, R. Bucala, Migration inhibitory factorexpression in experimentally induced endotoxemia, Am. J. Pathol. 150 (1997)235–246.
[3] J.A. Baugh, R. Bucala, Macrophage migration inhibitory factor, Crit. Care. Med.30 (2002) S27–S35.
[4] B. Beutler, Innate immunity: an overview, Mol. Immunol. 40 (2004) 845–859.[5] S. Bird, J. Zou, T. Wang, B. Munday, C. Cunningham, C.J. Secombes, Evolution of
interleukin-1beta, Cytokine Growth Factor Rev. 13 (2002) 483–502.[6] P.D. Bloom, E.C. Boedecker, Mucosal immune responses to intestinal bacterial
pathogens, Semin. Gastrointest. Dis. 7 (1996) 151–166.

460 S. Jantra et al. / General and Comparative Endocrinology 173 (2011) 454–460
[7] E. Cario, G. Gerken, D.K. Podolsky, ‘‘For whom the best tolls!’’ – innate defensemechanisms and survival strategies of the intestinal epithelium againstluminal pathogens, Z Gastroenterol. 40 (2002) 983–990.
[8] C. Cateni, L. Paulesu, E. Bigliardi, W.C. Hamlett, The interleukin 1 (IL-1) systemin the uteroplacental complex of a cartilaginous fish, the smoothhound shark,Mustelus canis, Reprod. Biol. Endocrinol. 1 (2003) 25.
[9] F. Colotta, F. Re, M. Muzio, R. Bertini, N. Polentarutti, M. Sironi, J.G. Giri, S.K.Dower, J.E. Sims, A. Mantovani, Interleukin-1 type II receptor: a decoy targetfor IL-1 that is regulated by IL-4, Science 261 (1993) 472–475.
[10] C.A. Dinarello, Interleukin-1, Cytokine Growth Factor Rev. 8 (1997) 253–265.[11] C.A. Dinarello, IL-1: discoveries, controversies and future directions, Eur. J.
Immunol. 40 (2010) 595–653.[12] X. Du, A. Poltorak, Y. Wei, B. Beutler, Three novel mammalian toll-like
receptors: gene structure, expression, and evolution, Eur. Cytokine Network 11(2000) 362–371.
[13] S.P. Eisenberg, M.T. Brewer, E. Verderber, P. Heindal, R.C. Thompson,Interleukin 1 receptor antagonist is a member of the interleukin 1 genefamily: evolution of a cytokine control mechanism, Proc. Natl. Acad. USA 88(1991) 5232–5236.
[14] J.V. Fahey, T.M. Schaefer, J.Y. Channon, C.R. Wira, Secretion of cytokines andchemokines by polarized human epithelial cells from the female reproductivetract, Hum. Reprod. 20 (2005) 1439–1446.
[15] P.G. Fallon, R.L. Allen, T. Rich, Primitive toll signaling: bugs, flies, worms andman, Trends Immunol. 22 (2001) 63–66.
[16] F.P. Faraone, F. Lillo, G. Giacalone, M. Lo Valvo, The large invasive population ofX. laevis in Sicily, Italy, Amphibia–Reptilia 29 (2008) 405–412.
[17] M.R. Faria, M.S. Hoshida, E.A. Ferro, F. Ietta, L. Paulesu, E. Bevilacqua,Spatiotemporal patterns of macrophage migration inhibitory factor (Mif)expression in the mouse placenta, Reprod. Biol. Endocrinol. 8 (2010) 95.
[18] G.R. Fingerle-Rowson, R. Bucala, Neuroendocrine properties of macrophagemigration inhibitory factor (MIF), Immunol. Cell. Biol. 79 (2001) 368–375.
[19] H. Flaster, J. Bernhagen, T. Calandra, R. Bucala, The macrophage migrationinhibitory factor-glucocorticoid dyad: regulation of inflammation andimmunity, Mol. Endocrinol. 21 (2007) 1267–1280.
[20] R.D. Fusunyan, N.N. Nanthakumar, M.E. Baldeon, W.A. Walker, Evidence for ainnate immune response I the immature human intestine: toll-like receptorson fetal eterocytes, Pediatric. Res. 49 (2001) 589–593.
[21] Y. Gon, Toll-like receptors and airway inflammation, Allergol. Int. 57 (2008)33–37.
[22] K.M. Hart, A.S. Murphy, K.T. Barrett, C.R. Wira, P.M. Guyre, P.A. Pioli, Functionalexpression of pattern recognition receptors in tissues of the human femalereproductive tract, J. Reprod. Immunol. 80 (2009) 33–40.
[23] J.A. Hoffmann, J.M. Reichhart, Drosophila innate immunity: an evolutionaryperspective, Nat. Immunol. 3 (2002) 121–126.
[24] F. Ietta, Y. Wu, R. Romagnoli, N. Soleymanlou, B. Orsini, S. Zamudio, L. Paulesu,I. Caniggia, Oxygen regulation of macrophage migration inhibitory factor inhuman placenta, Am. J. Physiol. Endocrinol. Metab. 292 (2007) E272–280.
[25] Y. Ikoma, S. Nomura, T. Ito, Y. Katsumata, M. Nakata, K. Iwanaga, M. Okada, F.Kikkawa, K. Tamakoshi, T. Nagasaka, M. Tsujimoto, S. Mizutani, Interleukin-1bstimulates placental leucine aminopeptidase/oxytocinase expression in BeWochoriocarcinoma cells, Mol. Hum. Reprod. 9 (2003) 103–110.
[26] A. Ishii, M. Kawasaki, M. Matsumoto, S. Tochinai, T. Seya, Phylogenetic andexpression analysis of amphibian Xenopus toll-like receptors, Immunogenetics59 (2007) 281–293.
[27] S. Jantra, E. Bigliardi, R. Brizzi, F. Ietta, N. Bechi, L. Paulesu, Interleukin 1 inoviductal tissues of viviparous, oviparous, and ovuliparous species ofamphibians, Biol. Reprod. 76 (2007) 1009–1015.
[28] A.M. Jelaso, S. Acevedo, T. Dang, A. Lepere, C.F. Ide, Interleukin 1 beta and itstype 1 receptor are expressed in developing neural circuits in the frog, X. laevis,J. Comp. Neurol. 394 (1998) 242–251.
[29] R. Kleemann, A. Hausser, G. Geiger, R. Mischke, A. Burger-Kentischer, O. Flieger,F.J. Johannes, T. Roger, T. Calandra, A. Kapurniotu, M. Grell, D. Finkelmeier, H.Brunner, J. Bernhagen, Intracellular action of the cytokine MIF to modulate AP-1 activity and the cell cycle through Jab1, Nature 408 (2000) 211–216.
[30] H. Lue, R. Kleemann, T. Calandra, T. Roger, J. Bernhagen, Macrophage migrationinhibitory factor (MIF): mechanisms of action and role in disease, MicrobesInfect. 4 (2002) 449–460.
[31] C. Luo, L. Zheng, Independent evolution of toll and related genes in insects andmammals, Immunogenetics 51 (2000) 92–98.
[32] M.U. Martin, W. Falk, The interleukin-1 receptor complex and interleukin-1signal transduction, Eur. Cytokine Netw. 8 (1997) 5–17.
[33] R. Medzhitov, C. Janeway, The Toll receptor family and microbial recognition,Trends Microbiol. 8 (2000) 452–456.
[34] R. Medzhitov, P. Preston-Hurlburt, C.A. Janeway, A human homologue of theDrosophila Toll protein signals activation of adaptive immunity, Nature 388(1997) 394–397.
[35] R.A. Mitchell, R. Bucala, MIF signal transduction initiated by binding to CD74, J.Exp. Med. 197 (2003) 1467–1476.
[36] A. Muir, G. Soong, S. Sokol, B. Reddy, M.I. Gomez, von A. Heeckeren, A. Prince,Toll-like receptors in normal and cystic fibrosis airway epithelial cells, Am. J.Respir. Cell. Mol. Biol. 30 (2004) 777–783.
[37] L. Paulesu, J. Bhattacharjee, N. Bechi, R. Romagnoli, S. Jantra, F. Ietta, Pro-inflammatory cytokines in animal and human gestation, Curr. Pharm Des. 16(2010) 3601–3615.
[38] L. Paulesu, E. Bigliardi, E. Paccagnini, F. Ietta, C. Cateni, C.P. Guillaume, B.Heulin, Cytokines in the oviparity/viviparity transition: evidence of theinterleukin-1 system in a species with reproductive bimodality, the lizardLacerta vivipara, Evol. Dev. 7 (2005) 282–288.
[39] L. Paulesu, C. Cateni, R. Romagnoli, F. Ietta, V. Dantzer, Variation in macrophagemigration-inhibitory factor immunoreactivity during porcine gestation, BiolReprod. 72 (2004) 949–953.
[40] L. Paulesu, S. Jantra, F. Ietta, R. Brizzi, E. Bigliardi, Interleukin-1 in reproductivestrategies, Evol. Dev. 10 (2008) 778–788.
[41] L. Paulesu, R. Romagnoli, M. Marchetti, M. Cintorino, P. Ghiara, F.M. Guarino, G.Ghiara, Cytokines in the viviparous reproduction of squamate reptiles:interleukin-1 alpha (IL-1 alpha) and IL-1 beta in placental structures of askink, Placenta 16 (1995) 193–205.
[42] P.A. Pioli, E. Amiel, T.M. Schaefer, J.E. Connolly, C.R. Wira, P.M. Guyre,Differential expression of toll-like receptors 2 and 4 in tissues of the humanfemale reproductive tract, Infect. Immun. 72 (2004) 5799–5806.
[43] A.J. Quale, The innate and early immune response to pathogen challenge in thefemale genital tract and the pivotal role of epithelial cells, J. Reprod. Immunol.57 (2002) 61–79.
[44] J.C. Roach, G. Glusman, L. Rowen, A. Kaur, M.K. Purcell, K.D. Smith, L.E. Hood, A.Aderem, The evolution of vertebrate toll-like receptors, Proc. Natl. Acad. Sci.USA 102 (2005) 9577–9582.
[45] T. Roger, J. David, MP. Glauser, T. Calandra, MIF regulates innate immuneresponses through modulation of toll-like receptor 4, Nature 414 (2001) 920–924.
[46] T. Roger, C. Froideyaux, C. Martin, T. Calandra, Macrophage migrationinhibitory factor (MIF) regulates host responses to endotoxin throughmodulation of toll-like receptor 4 (TLR4), J. Endotoxin. Res. 9 (2003) 119–123.
[47] J.E. Sims, M.A. Gayle, J.L. Slack, M.R. Alderson, T.A. Bird, J.G. Giri, F. Colotta, F.Re, A. Mantovani, K. Shanebeck, Interleukin 1 signaling occurs exclusively viathe type I receptor, Proc. Natl. Acad. Sci. USA 90 (1993) 6155–6159.
[48] M. Suzuki, Y. Takamura, M. Maéno, S. Tochinai, D. Iyaguchi, I. Tanaka, J.Nishihira, T. Ishibashi, X. laevis macrophage migration inhibitory factor isessential for axis formation and neural development, J. Biol. Chem. 279 (2004)21406–21414.
[49] B.D. Todd, Parasite lost? An overlooked hypothesis for the evolution ofalternative reproductive strategies in amphibians, Am. Nat. 170 (2007) 793–799.
[50] J. Vilcek, J. Le, Immunology of cytokines: an introduction, in: A. Thomson (Ed.),The cytokine Handbook, Academic Press, London, 1994, pp. 1–19.
[51] G.W. Warr, R.W. Chapman, L.C. Smith, Evolutionary immunobiology: newapproaches, new paradigms, Dev. Comp. Immunol. 27 (2003) 257–262.
[52] C.R. Wira, K.S. Grant-Tschudy, M.A. Crane-Godreau, Epithelial cells in thefemale reproductive tract: a central role as sentinels of immune protection,Am. J. Reprod. Immunol. 53 (2005) 65–76.
[53] N. Yoshizaki, Fine structure of oviducal epithelium of X. laevis in relation to itsrole in secreting egg envelopes, J. Morphology 184 (1985) 155–169.
[54] J. Zou, R.M. Bird, J. Horton, C. Cunningham, C.J. Secombes, Molecular cloning ofthe gene for interleukin-1b from X. laevis and analysis of expression in vivo andin vitro, Immunogenetics 51 (2000) 332–338.
Related Documents











