The new england journal of medicine n engl j med 368;6 nejm.org february 7, 2013 543 brief report Brain Dopamine–Serotonin Vesicular Transport Disease and Its Treatment Jennifer J. Rilstone, B.Sc., Reem A. Alkhater, M.D., and Berge A. Minassian, M.D. From the Institute of Medical Science, University of Toronto (J.J.R., R.A.A., B.A.M.), and the Program in Genetics and Genome Biology (J.J.R., B.A.M.) and Division of Neurology, Department of Pediatrics (R.A.A., B.A.M.), Hospital for Sick Chil- dren — both in Toronto. Address reprint requests to Dr. Minassian or Dr. Alkhater at Rm. 6536B, Hospital for Sick Children, 555 University Ave., Toronto, ON M5G 1X8, Canada, or at [email protected] or [email protected]. Ms. Rilstone and Dr. Alkhater contributed equally to this article. This article was published on January 30, 2013, at NEJM.org. N Engl J Med 2013;368:543-50. DOI: 10.1056/NEJMoa1207281 Copyright © 2013 Massachusetts Medical Society. SUMMARY We describe a disease encompassing infantile-onset movement disorder (including severe parkinsonism and nonambulation), mood disturbance, autonomic instabil- ity, and developmental delay, and we describe evidence supporting its causation by a mutation in SLC18A2 (which encodes vesicular monoamine transporter 2 [VMAT2]). VMAT2 translocates dopamine and serotonin into synaptic vesicles and is essential for motor control, stable mood, and autonomic function. Treatment with levodopa was associated with worsening, whereas treatment with direct dopamine agonists was followed by immediate ambulation, near-complete correction of the movement disorder, and resumption of development. K nown disorders of biogenic amine neuromediators (dopamine, norepinephrine, epinephrine, and serotonin) involve defects in nine en- zymes 1-9 and one transporter. 10 Affected persons present in early childhood with symptoms referable to the affected neurotransmitter, and the disorders are diagnosed by measurement of neurotransmitter breakdown products in the cere- brospinal fluid (CSF). A deficiency in dopamine results in movement disorder; de- ficient norepinephrine or epinephrine causes autonomic dysfunction; and serotonin deficiency leads to sleep and psychiatric disturbances. 2,3,6 We describe members of a family with symptoms of deficiencies in dopamine (dystonia, parkinsonism, and oculogyric crises), serotonin (sleep and mood distur- bance), and epinephrine and norepinephrine (diaphoresis, temperature instability, ptosis, and postural hypotension), with no demonstrable deficiency of neurotrans- mitters in the CSF. Genome investigation revealed a mutation in the gene encoding VMAT2 that compromises transport of biogenic amines into synaptic vesicles, resulting in impairment of their synaptic transmission without detectable reduc- tions in their amounts. CASE REPORT Eight children of an extended consanguineous Saudi Arabian family had similar clinical symptoms of a complex movement disorder that was inherited in an auto- somal recessive fashion (Fig. 1A; and Table S1 in the Supplementary Appendix, available with the full text of this article at NEJM.org). The parents were unaffected, but at least five had clinical depression. The index patient, when she presented to us, was a 16-year-old girl with global developmental delay and abnormal movements. She had first been brought to medical attention at 4 months of age with hypotonia, loss of acquired head con- trol, and paroxysmal stereotyped episodes of persistent eye deviation and crying The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission. Copyright © 2013 Massachusetts Medical Society. All rights reserved.

Brain Dopamine–Serotonin Vesicular Transport Disease and Its Treatment.pdf
Nov 01, 2014
Brain Dopamine–Serotonin Vesicular Transport Disease and Its Treatment
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

T h e n e w e ngl a nd j o u r na l o f m e dic i n e
n engl j med 368;6 nejm.org february 7, 2013 543
brief report
Brain Dopamine–Serotonin Vesicular Transport Disease and Its Treatment
Jennifer J. Rilstone, B.Sc., Reem A. Alkhater, M.D., and Berge A. Minassian, M.D.
From the Institute of Medical Science, University of Toronto (J.J.R., R.A.A., B.A.M.), and the Program in Genetics and Genome Biology (J.J.R., B.A.M.) and Division of Neurology, Department of Pediatrics (R.A.A., B.A.M.), Hospital for Sick Chil-dren — both in Toronto. Address reprint requests to Dr. Minassian or Dr. Alkhater at Rm. 6536B, Hospital for Sick Children, 555 University Ave., Toronto, ON M5G 1X8, Canada, or at [email protected] or [email protected].
Ms. Rilstone and Dr. Alkhater contributed equally to this article.
This article was published on January 30, 2013, at NEJM.org.
N Engl J Med 2013;368:543-50.DOI: 10.1056/NEJMoa1207281Copyright © 2013 Massachusetts Medical Society.
SUMM A R Y
We describe a disease encompassing infantile-onset movement disorder (including severe parkinsonism and nonambulation), mood disturbance, autonomic instabil-ity, and developmental delay, and we describe evidence supporting its causation by a mutation in SLC18A2 (which encodes vesicular monoamine transporter 2 [VMAT2]). VMAT2 translocates dopamine and serotonin into synaptic vesicles and is essential for motor control, stable mood, and autonomic function. Treatment with levodopa was associated with worsening, whereas treatment with direct dopamine agonists was followed by immediate ambulation, near-complete correction of the movement disorder, and resumption of development.
Known disorders of biogenic amine neuromediators (dopamine, norepinephrine, epinephrine, and serotonin) involve defects in nine en-zymes1-9 and one transporter.10 Affected persons present in early childhood
with symptoms referable to the affected neurotransmitter, and the disorders are diagnosed by measurement of neurotransmitter breakdown products in the cere-brospinal fluid (CSF). A deficiency in dopamine results in movement disorder; de-ficient norepinephrine or epinephrine causes autonomic dysfunction; and serotonin deficiency leads to sleep and psychiatric disturbances.2,3,6
We describe members of a family with symptoms of deficiencies in dopamine (dystonia, parkinsonism, and oculogyric crises), serotonin (sleep and mood distur-bance), and epinephrine and norepinephrine (diaphoresis, temperature instability, ptosis, and postural hypotension), with no demonstrable deficiency of neurotrans-mitters in the CSF. Genome investigation revealed a mutation in the gene encoding VMAT2 that compromises transport of biogenic amines into synaptic vesicles, resulting in impairment of their synaptic transmission without detectable reduc-tions in their amounts.
C A SE R EPORT
Eight children of an extended consanguineous Saudi Arabian family had similar clinical symptoms of a complex movement disorder that was inherited in an auto-somal recessive fashion (Fig. 1A; and Table S1 in the Supplementary Appendix, available with the full text of this article at NEJM.org). The parents were unaffected, but at least five had clinical depression.
The index patient, when she presented to us, was a 16-year-old girl with global developmental delay and abnormal movements. She had first been brought to medical attention at 4 months of age with hypotonia, loss of acquired head con-trol, and paroxysmal stereotyped episodes of persistent eye deviation and crying
The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission.
Copyright © 2013 Massachusetts Medical Society. All rights reserved.

T h e n e w e ngl a nd j o u r na l o f m e dic i n e
n engl j med 368;6 nejm.org february 7, 2013544
B
D E
C
A
5
4 2
6 4 3
I-1
II-1
II-7 II-8
III-1
IV-1
V-1 V-2 V-3 V-4 V-5 V-6 V-7 V-8 V-9 V-10 V-11 V-12 V-13 IV-8 IV-9 IV-10
VI-4VI-3VI-2VI-1
IV-2 IV-3 IV-4 IV-5 IV-6 IV-7
III-2 III-3 III-4 III-8 III-11
III-5 III-6 III-7 III-9 III-10
II-2 II-3 II-4 II-5 II-6
I-2
RS4751923RS2907578RS4751940RS7071452RS3781523RS10787635RS1077114RS12245521RS2915753RS733216RS1615201RS1638438RS181499RS181586RS703419RS856003RS912745RS1925251RS10510038RS11198226RS10886215RS4411217RS1766253RS10886296RS7091672RS943269RS2297696RS506657RS915120RS7071853RS196272RS112029RS2273747
117431164117550674117703211117811340117875250117937271118004195118287023118354832118441800118515206118919973119173414119258198119324784119393553119488150119555244119638844119755841120012268120212447120338216120512222120586411120705108120906457121127172121180103121301596121384677121430518121581501
GAAAGAAGAGCGAAGAAGAAGAGAAGAAGGGAG
AAGGAAAGAGCGAAGAAGAAGAGAAGAAGGGAG
GGGAGCGGGACACGGAGAAAGAAAGGGGGGAGG
AAGGAAAGAGCGAAGAAGAAGAGAAGAAGGGAG
GGAGGCGAGAAACGAAGAGGAGAGGGGGAAAGA
GAGGAAGGAGCGCAGAGGAAGAAAAGGGGGAAG
GAAAGAAGAGCGAAGAAGAAGAGAAGAAGGGAG
AAGGAAAGAGCGAAGAAGAAGAGAAGAAGGGAG
GAAAGAAGAGCGAAGAAGAAGAGAAGAAGGGAG
AAGGAAAGAGCGAAGAAGAAGAGAAGAAGGGAG
AAGAAAAGAGCGAAGAAGAAGAGAAGAAGGGAG
AAGAAAAGAGCGAAGAAGAAGAGAAGAAGGGAG
AAGAGCGGGGAACAGGGGGGGAAAGGAGGAAGA
AAGGAAAGAGCGAAGAAGAAGAGAAGAAGGGAG
GAAAGAAGAGCGAAGAAGAAGAGAAGAGAAAAG
AAGGAAAGAGCGAAGAAGAAGAGAAGAAGGGAG
TM9VMAT2 TM10HomoPatientPanMacacaMusRattusCanisBosMonodelphisGallusTetraodonDanioDrosophilaC. eleganshVMAT1
ILCIPFAKNIYGLIAPNFGVGFAIGMVDSSILCIPFAKNIYGLIALNFGVGFAIGMVDSSILCIPFAKNIYGLIAPNFGVGFAIGMVDSSILCIPFAKNIYGLIAPNFGVGFAIGMVDSSILCIPFAKNIYGLIAPNFGVGFAIGMVDSSILCIPFAKNIYGLIAPNFGVGFAIGMVDSSILCIPFAKNIYGLIAPNFGVGFAIGMVDSSILCIPLAKNIYGLIAPNFGVGFAIGMVDSSVLCVPLARNIYGLIAPNFGIGFAIGMVDSSILCVPFAKNIYGLIAPNFGVGFAIGMVDSSVICVPFATSIYGLILPNFGVGFAIGMVDSSILCVPLAKDIYGLIVPNFGVGFAIGMVDSSLIFIPMATSITHLIIPNAGLGFAIGMVDSSLLSIPSATSVAGLIIPHALLGFSIGMIDASLLCVPLAHNIFGLIGPNAGLGLAIGMVDSS
V-6 V-7 V-8 V-9 V-10 V-3 V-4 VI-2 G C T C C G A A C
Normal
Parent (IV-3)
Patient (V-6)c.1160C→T
*
MALS
SS
SYY
YY
Y
Y
Y
YY
Y
S
SS S
S
D
D
DD
D
D
DD
D D
D
D D
SF
F
F F
S
SSC
C
S
S
S
S
S
SS
G
GIP
P
P
PPP
P
P
PP
PP
PP
P
P
P
P
I
II
II
I
II
I
I
I
II
II
I
GG
G
G
G
G
G
G
S
S
C
S S
LE
E
EH
H
H
H
H
H
H
ET
T
T
TT
T
TT
TT
T
TT
T
T
T
T
T
E
E
E E
E
E
E
E
EE
EEEE
LVR
W
W
RRRK
K
KN
NN
N
K
K
K
K
K K K
K
K
KK
R
R
R
R
R RR
RN
N
NN
N
N
NN
N
R R
V
V
V
V
V
V
V V V
V
LQ
Q
Q
Q
Q
Q Q
Q
Q
Q
LL
L
LL
L
L
LL
L
L
L
L
A
A
A
A
A
AA
A
A AA
A
A
M
M
MMM
M
M
MW
F
P
P
P
P
P
PPP
P
P F
FF
FFF
F
FF
F
FF
FF
F
F
F
F
FG G
G
GG
GG
GGG
GGQ
Q
Q
G
G
G G
GG
G
GG
G GG
G G
G
G
G
G
FF
F
AC
CY
Y
Y
Y
Y
Y
Y C
C
C
C
C
L
LD
D
DD
D
L
LL
LLL
L
W
W
L
L
L
L
L
LL
L
LLL
L
L
L
LLLL
LL
L
LL
LL II I
I
I
I
II
II
I
II
I
III
I I I
II
I
II
I
IIT
T
T
TT
T
T
T
III
IL
L
L L
L
LL
L
A
A
AVV
V V
VVVV
V
V
VV
V
V
VV
V
V
VVVV
VA
AA
A
A
A
A
A
A
A
A
A
A
A
AA
AA
AA
A
A
AA
A
AKS S SS SS S
S
S
S SS S
S S
SS
S
A
A
A
MM
MM
M
M
MM
M
MM
M
MN
N
N
N
M M
M
MCytoplasm
Vesicle Lumen
The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission.
Copyright © 2013 Massachusetts Medical Society. All rights reserved.

brief report
n engl j med 368;6 nejm.org february 7, 2013 545
lasting hours. Video electroencephalographic monitoring had ruled out seizures, and a symp-tom-based diagnosis of oculogyric crisis had been made. Development had been normal ini-tially but had slowed after presentation. The girl sat at 30 months, crawled at 4 years, and walked at 13 years.
At 16 years of age, she had fatigue, excessive diaphoresis, profuse nasal and oropharyngeal secretions, noisy breathing, hypernasal speech, poor distal perfusion, cold hands and feet, dis-rupted sleep, hypotonia, dysarthria, and ataxia. There was no diurnal variation and no improve-ment with administration of vitamin B6 or folinic acid. Neurologic examination revealed ptosis, hy-pomimia, facial dyskinesia, and limited upward gaze. She had axial hypotonia and appendicular hypertonia specifically involving extensor mus-cles of the arms and legs. Her deep-tendon re-flexes were minimally hyperreflexic, and her plantar responses were flexor. Coordination test-ing revealed a fine tremor and dysdiadochokine-sia in the arms and legs. The patient’s gait was parkinsonian, with typical shuffling; her posture was stooped, and her postural reflexes were di-minished. She walked with bilateral alternating dystonia of the hands and feet, with intermittent toe walking and foot inversion, and was unable to perform a tandem walk.
Results of basic blood tests, metabolic screen-ing tests (Table S2 in the Supplementary Ap-pendix), repeat video electroencephalography, magnetic resonance imaging, and magnetic res-onance spectroscopy were normal. A lumbar
puncture was performed in a 2-year-old affected sibling, and analysis of the CSF showed normal levels of neurotransmitter metabolites, intermedi-ates, and precursors (Table S3 in the Supplemen-tary Appendix). However, urinary neurotransmit-ter testing revealed elevated levels of monoamine metabolites (5-hydroxyindoleacetic acid, 17.6 μg per deciliter [reference range, 0 to 6.0]; homo-vanillic acid, 14.1 μg per milligram of urinary creatinine [reference range, 0 to 13.4]) and de-creased levels of measurable monoamines (nor-epinephrine, 1 μg per deciliter [reference range, 4 to 29]; dopamine, 19 μg per deciliter [refer-ence range, 40 to 260]) (Table S3 in the Supple-mentary Appendix).
On the basis of the parkinsonism and the diminished urinary dopamine, the proband and three younger affected siblings were given levo-dopa–carbidopa, which within 1 week resulted in major deterioration, with the appearance of intense chorea and worsened dystonia. Discon-tinuation of the medication led to a rapid return to baseline function in all four children.
Me thods
GENETIC STUDIES
The study was approved by the research ethics board of the Hospital for Sick Children, and par-ents provided written informed consent. We geno-typed more than 300,000 single-nucleotide poly-morphisms (SNPs) in eight family members, V-3, V-4, V-6, V-7, V-8, V-9, V-10, and VI-2 (Fig. 1A) (with the use of an Illumina 300K SNP microarray), followed by homozygosity mapping to identify the homozygous loci shared by the affected chil-dren. A subset of 2500 SNPs with a minimal al-lele frequency greater than 0.4 in the population genotyped as part of the international HapMap study and with average spacing of approximately 1.0 Mb was selected for parametric linkage analy-sis. We used Sanger sequencing of candidate gene exons to identify the mutation, whole-exome se-quencing (Agilent V4 50Mb capture kit and Illu-mina HiSeq 2000 sequencing) to rule out other mutations, and TaqMan genotyping to confirm the absence of the mutation in controls.
FUNCTIONAL ANALYSIS OF P387L-MUTANT VMAT2
We engineered a construct encoding VMAT2 that contained the P387L substitution, and we carried out an assay of vesicular serotonin uptake in a
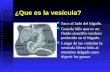
Figure 1 (facing page). The Causative Defect.
Panel A depicts the family pedigree. Squares denote male family members, circles female family members, black symbols affected family members, and diamonds miscarriages; double bars indicate consanguineous marriages, and the arrow indicates the index patient. Panel B shows the homozygous haplotype shared by the affected persons. Panel C shows electropherograms of a portion of SLC18A2 containing the mutation. Panel D depicts the vesicular monoamine transporter 2 (VMAT2) protein within the synaptic vesicle membrane; the arrow indicates the mutated proline. Panel E shows the pri-mary sequence of the portion of human VMAT2, which in our patients contains the mutated proline, and cor-responding sequences in various orthologues and in the VMAT1 paralog of neuroendocrine cells. TM9 and TM10 are parts of the 9th and 10th transmembrane domains, respectively, of VMAT2. Panel D adapted from Erickson and Eiden.14
The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission.
Copyright © 2013 Massachusetts Medical Society. All rights reserved.

T h e n e w e ngl a nd j o u r na l o f m e dic i n e
n engl j med 368;6 nejm.org february 7, 2013546
heterologous cell system (see the Methods sec-tion in the Supplementary Appendix). Transport mediated by VMAT2 was measured by incubating membrane preparations with tritiated serotonin, followed by rapid washing and filtration to retain vesicles with trapped substrate.
R esult s
MUTATION IDENTIFICATION
Homozygosity mapping identified a single homo-zygous 3.2-Mb interval on chromosome 10q in the region of 10q25.3-26.11 that was shared by five affected family members but not by unaf-fected members (Fig. 1B). Parametric linkage analysis revealed a significant logarithm of odds (lod) score of 4.1 in this region. Another locus, on chromosome 3, yielded a significant lod score of 3.1 but did not correspond to a region of shared homozygosity. We sequenced exons and exon–intron boundaries of eight genes known to have neuronal functions and observed a novel variant (c.1160C→T) in exon 13 (Fig. 1C), which is predicted to result in a substitution of leucine for proline at position 387 (p.P387L) in VMAT2. The variant was homozygous in affected family mem-bers but not in 78 unaffected members, 26 of whom carried the variant in the heterozygous state.
We also performed whole-exome sequencing in the proband, which independently identified the SLC18A2 change and revealed no other novel nonsynonymous variant in the linked region of shared homozygosity. SLC18A2 c.1160C→T is not present in data sets of sequenced genomes, in-cluding the 1000-genome database. In addition, screening for SLC18A2, one of the most exten-sively studied candidate genes for involvement in Parkinson’s disease, was previously performed in 704 healthy persons of diverse ethnic back-grounds and 452 patients with Parkinson’s dis-ease,11-13 none of whom had the c.1160C→T change. Collectively, these results suggest that SLC18A2 c.1160C→T is the causative defect in this family.
FUNCTIONAL CHARACTERIZATION OF P387L-MUTANT VMAT2
SLC18A2 encodes the VMAT2 protein located in membranes of monoamine synaptic vesicles (Fig. 1D, 2A, and 2B). Proline residues adjacent to
transmembrane segments have major structural effects and are overrepresented among residues subject to disease-causing substitutions.15 The proline residue in the 387 position (Pro387) of the VMAT2 protein is adjacent to a transmem-brane segment (Fig. 1D). Sequence alignment shows that Pro387 is highly conserved through evolution and thus suggests that its substitution is likely to be deleterious. It is also conserved in the paralogous protein VMAT1 and in the Cae-norhabditis elegans CAT-1 protein — the single ve-sicular monoamine transporter in nematodes (Fig. 1E).16 The residue is not conserved in the vesicular acetylcholine transporter, which main-tains 39% identity with VMAT2; this finding im-plies that Pro387 may have a specific role in monoamine transport.
To determine the effect of the P387L muta-tion on VMAT2 transport activity, we transiently and separately expressed nonmutant and mutant human VMAT2 in COS-7 cells. Immunoblot analy-sis of membrane preparations confirmed equiva-lent levels of mature glycosylated VMAT2 in parallel transfections, suggesting that there was no major defect in protein processing. However, P387L-mutant VMAT2 showed dramatically de-creased activity as compared with nonmutant VMAT2 (Fig. 2C). Use of the specific VMAT in-hibitor reserpine confirmed that P387L-mutant VMAT2 still exhibited some weakly measurable uptake (Fig. 2D). Thus, the P387L mutation results in severe, but not complete, loss of function.
TREATMENT
Defective monoamine loading into synaptic vesi-cles, and therefore neurotransmission, was con-sistent with symptoms of monoamine deficiency in affected members of the family, despite their normal levels of brain monoamine. With this in-sight, we gave the proband a direct dopamine-receptor agonist (pramipexole), which resulted, within 1 week, in dramatic and sustained disap-pearance of parkinsonism and dystonic attacks and improvement in other symptoms (Table 1). We then provided treatment to the younger sib-lings, who also had improvement. It seemed that the younger the affected child, the more substan-tial the recovery (Table 1). The affected children are now in their 32nd month of treatment, with continuing benefits and minimal side effects (slight overactivity and weight loss).
The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission.
Copyright © 2013 Massachusetts Medical Society. All rights reserved.

brief report
n engl j med 368;6 nejm.org february 7, 2013 547
DISCUSSION
The mutation in SLC18A2 that we describe here is expected to affect monoamine neurotransmis-sion and thus result in a phenotype that has over-
lap with all monoamine disorders. Because move-ment disorder is conspicuous among symptoms of monoamine disturbance, the clinical picture of the disease that we describe is closest to that of diseases affecting dopamine — chiefly, deficien-
Dopamine
Epinephrine
Dopamine
NE
DβH
PNMT
MHPG
VMA
HVA
Serotonin
Serotonin
5-HTP
5-HIAA
TryptophanPresynaptic
neuron
Postsynaptic neuron
Glia
Dopaminetransporter
Monoaminereceptor
Tryptophan hydroxylase
L-DOPA
Tyrosine
Tyrosinehydroxylase
VMAT2
VMAT2
MAOCOMT
MAOCOMT
AADC
A B
C D
MAOCOMT
MAOCOMT
VMAT2
Sero
toni
n Tr
ansp
ort (
pmol
/mg)
25
20
15
5
10
00 5 10 15 20
− Reserpine+ Reserpine
Wild-type VMAT2P387L-mutant VMAT2Control
Minutes
VMAT2
GAPDH
~55kD
Wild
-type
P387L
Vecto
r
~45kDSe
roto
nin
Tran
spor
t (pm
ol/m
g)
0
5
10
15
20
25
30
Wild-typeVMAT2
P387L-mutantVMAT2
3-OMD
qBH2
BH4
SR
Figure 2. Monoamine Metabolism, Vesicular Loading, Synaptic Transmission, and the Loading Defect in the Patients.
Panel A summarizes the metabolism of monoamines and their transport into synaptic vesicles; norepinephrine (NE) and epinephrine are synthesized from dopamine within the vesicle. Panel B depicts monoamine neurotransmission; the site of action of the dopamine agonist in the patients described here is at the postsynaptic neuron membrane. Panel C shows the time course of tritiated serotonin uptake into vesicles prepared from COS-7 cells transfected with wild-type or mutant (P387L) human VMAT2 or with the vector alone (control). Western blot analy-sis showed equivalent VMAT2 levels and post-translational processing in assay lysates. Panel D shows the uptake of tritiated serotonin (over a period of 10 minutes) by vesicles prepared from cells transfected with wild-type or P387L-mutant VMAT2, with and without the addition of 10 μM of the specific VMAT inhibitor reserpine. AADC denotes aromatic l-amino acid decarboxylase, BH4 tetrahydrobiopterin, COMT catechol-O-methyltransferase, DβH dopamine β-hydroxylase, GAPDH glyceraldehyde-3-phophate dehydrogenase, HIAA hydroxyindoleacetic acid, HTP hydroxytryptophan, HVA homovanillic acid, MAO monoamine oxidase, MHPG 3-methoxy-4-hydroxyphenylglycol, OMD O-methyldopa, PNMT phenylethanolamine N-methyltransferase, qBH2 quinonoid dihydrobiopterin, SR sepiapterin reductase, and VMA vanillylmandelic acid.
The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission.
Copyright © 2013 Massachusetts Medical Society. All rights reserved.

T h e n e w e ngl a nd j o u r na l o f m e dic i n e
n engl j med 368;6 nejm.org february 7, 2013548
cies in dopamine transporter, tetrahydrobiopterin, tyrosine hydroxylase, and aromatic l-amino acid decarboxylase (AADC) (Fig. 2A and 2B). The phe-notype of the affected siblings has particular similarity to AADC deficiency in that it improves with direct dopamine agonism but not with levo-dopa, although the siblings had greater improve-ment than that typically observed in those with AADC deficiency treated with dopamine ago-nists, and rather than having a lack of response to levodopa, the siblings had a worsening of symptoms. Two other features that distinguish the disease we describe here from AADC defi-ciency are the lack of improvement with the AADC enzyme cofactor vitamin B6 and the ab-sence of worsening in the evening, which in AADC deficiency is the result of neurotransmit-ter depletion due to insufficient production.1,4,7,8
The standard diagnostic test in patients with suspected diseases of monoamine metabolism is the measurement of monoamine metabolites in the CSF. Because each specific defect results in a particular metabolite profile, this single test specifies the disease.2,3,6 Analysis of monoamines or their metabolites in urine is not reliable for the diagnosis of monoamine neurotransmitter diseases,2,3,6 except for one — AADC deficiency — in which increased 3-O-methyldopa with de-creased vanillylmandelic acid (Fig. 2A) in the proper clinical context is highly suggestive and generally confirmed by mutation analysis.1,4,7,8 In the present condition, the urine shows abnor-malities because VMAT2 also functions at sites outside the central nervous system, including the peripheral nervous system, adrenal medulla, and platelets.17 Our detection of abnormalities in the urine but not the CSF may reflect differ-ences in monoamine and metabolite stabilities, processing, and normal ranges between the brain and the periphery. In any case, it appears that AADC and VMAT2 deficiencies, which are meta-bolically and clinically similar disorders, could be screened for by urine testing and then con-firmed by gene sequencing, thus obviating the need for a lumbar puncture.
Direct characterization of the mutant VMAT2 protein in this study revealed a severe detriment of vesicular transporter function, which could be due to poor incorporation of the transporter into vesicle membranes or to loss of activity. Proline-to-leucine substitutions are generally considered to be deleterious to organismal fitness,18,19 on Ta
ble
1. R
elat
ions
hip
betw
een
Age
at I
nitia
tion
of D
opam
ine-
Ago
nist
The
rapy
and
Dis
ease
Cou
rse
in th
e Fo
ur A
ffec
ted
Sibl
ings
.
Var
iabl
eA
ge a
t In
itiat
ion
of T
hera
py
Patie
nt V
-6, 1
8 yr
Patie
nt V
-9, 1
1 yr
Patie
nt V
-10,
7 y
rPa
tient
V-1
1, 3
yr
Cog
nitio
n an
d ab
ility
to
lear
nM
ildly
impr
oved
Mild
ly im
prov
edM
oder
atel
y im
prov
edG
reat
ly im
prov
ed; p
atie
nt a
ble
to m
ake
stor
ies
from
pic
ture
s
Ocu
logy
ric
cris
esN
o fu
rthe
r ev
ents
; pat
ient
req
uire
d hi
gher
dos
e by
wei
ght t
han
her
sibl
ings
No
furt
her
even
tsN
o fu
rthe
r ev
ents
No
furt
her
even
ts
Dys
toni
aG
ait d
ysto
nia
pers
iste
dG
ait d
ysto
nia
pers
iste
dG
ait d
ysto
nia
pers
iste
dG
ait d
ysto
nia
impr
oved
Park
inso
nism
Impr
oved
Impr
oved
Impr
oved
Impr
oved
Fine
mot
or s
kills
Impr
oved
coo
rdin
atio
n; p
atie
nt a
ble
to fe
ed s
elf,
drin
k fr
om c
up, a
nd
hold
a p
en; i
mpr
oved
han
dwri
ting
Impr
oved
coo
rdin
atio
n; p
atie
nt le
arn-
ing
to h
old
a pe
n, u
nabl
e to
wri
te
or r
ead
Patie
nt le
arni
ng to
hol
d a
pen
and
drin
k fr
om a
cup
inde
pend
ently
, un
able
to w
rite
or
read
Patie
nt a
ble
to w
rite
, lea
rnin
g to
rea
d
Lang
uage
and
spe
ech
Dys
arth
ria
No
lang
uage
dev
elop
men
tPa
tient
cou
ld s
ay “
Mam
a” a
nd “
Papa
”N
orm
al la
ngua
ge d
evel
opm
ent a
nd
mild
dys
arth
ria
Gai
tIm
prov
ed p
ostu
re a
nd r
educ
ed fa
tigue
(p
atie
nt h
ad s
tart
ed w
alki
ng a
t 13
yr
of a
ge)
Patie
nt s
tart
ed w
alki
ng w
ithin
day
s af
ter
trea
tmen
tPa
tient
sta
rted
wal
king
with
in d
ays
afte
r tr
eatm
ent
Patie
nt s
tart
ed w
alki
ng w
ithin
day
s af
ter
trea
tmen
t
The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission.
Copyright © 2013 Massachusetts Medical Society. All rights reserved.

brief report
n engl j med 368;6 nejm.org february 7, 2013 549
the basis of analyses of amino acid substitutions in evolutionarily conserved proteins, and to be damaging to protein function, owing to a physi-cochemical difference.20 Proline places unique constraints on the flexibility of the peptide back-bone, particularly with respect to insertions of adjacent transmembrane segments.15
A complete knockout of Slc18a2 in mice results in a lack of exocytotic monoamine neurotrans-mission; the mutant animals feed poorly and die within days after birth.21,22 By contrast, mice that express just 5% native Vmat2 levels live to adulthood and have minor age-related motor deficits over time.23 The phenotypic spectrum of Vmat2 deficiency in mice is therefore broad and consistent with a requirement for large de-creases in protein function to cause severe motor symptoms.
We found that the motor phenotype was cor-rectable and that the extent of correction ap-peared to depend on the stage of the disease. If true, this dependency could be due to irrevers-ibly perturbed reorganization of dopamine path-ways in brains subjected to chronic deficiencies in monoamine neurotransmission during active brain development. Although the improvement in the patients in this study was striking, it was not complete, probably because of monoamine deficiency during development and also because of ongoing deficiencies of the non-dopamine amines and impairment in regulated release of dopamine.
Heterozygous mice with a single Slc18a2 allele have no motor phenotype but do have a depres-sive behavioral phenotype.24 We found a very high rate of depression among the parents of our patients (all five of the five parents interviewed reported depression). This is also seen in parents
of patients with AADC deficiency and is thought to be caused by clinically significant reductions in serotonin in these persons with hemizygous defects in the serotonin pathway.1,4,7,8 To what extent mutations in the genes encoding AADC and VMAT2 may contribute to common depres-sion and its heritability remains to be seen.
The initial selection of treatment of the af-fected children on the basis of clinical pheno-type alone (parkinsonism) led to severe, imme-diate worsening of the movement disorder. This was probably caused by the known toxicity of elevated levels of dopamine, in particular to do-paminergic neurons.25 Subsequent identification of the underlying pathophysiology allowed the rational selection of an appropriate treatment. A related severe disorder, sepiapterin reductase deficiency (see Fig. 2A), was recently diagnosed by means of whole-genome sequencing in 14-year-old fraternal twins; this previously known dis-order had been undiagnosed (and therefore un-treated) for many years because of the difficulties in obtaining a precise diagnosis for rare diseases.26 Diagnosis allowed treatment of the disorder, which led to recovery in those children.
Supported by the Canadian Institutes of Health Research. Dr. Minassian holds the University of Toronto Michael Bahen Chair in Epilepsy Research. Ms. Rilstone was supported by the Vanier Canada Graduate Scholarship from the National Sciences and Engineering Research Council of Canada.
Disclosure forms provided by the authors are available with the full text of this article at NEJM.org.
We thank the patients and their extended family; Drs. Mo-hammad Sheikh and Mohammad Domaia for local care of af-fected family members; Drs. Robert Edwards and Thomas Hnasko at the University of California, San Francisco, for guidance with the uptake assay; Drs. Marie-Thérèse Berthier and Denys Cyr and the University of Sherbrooke Biochemical Genetics Service for CSF monoamine and monoamine metabolite measurements; Drs. Stephen Scherer and Christian Marshall and the McLaughlin Foundation for whole-exome sequencing; and summer students Tarek Abdelhalim and John Bilbily for their contributions.
References
1. Brun L, Ngu LH, Keng WT, et al. Clin-ical and biochemical features of aromatic L-amino acid decarboxylase deficiency. Neurology 2010;75:64-71. [Erratum, Neu-rology 2010;75:576.]2. Hyland K. Clinical utility of mono-amine neurotransmitter metabolite analy-sis in cerebrospinal f luid. Clin Chem 2008;54:633-41.3. Kurian MA, Gissen P, Smith M, Heales S Jr, Clayton PT. The monoamine neurotransmitter disorders: an expanding range of neurological syndromes. Lancet Neurol 2011;10:721-33.4. Lee HC, Lai CK, Yau KC, et al. Non-
invasive urinary screening for aromatic L-amino acid decarboxylase deficiency in high-prevalence areas: a pilot study. Clin Chim Acta 2012;413:126-30.5. Longo N. Disorders of biopterin me-tabolism. J Inherit Metab Dis 2009;32:333-42. [Erratum, J Inherit Metab Dis 2009; 32:457.]6. Pons R. The phenotypic spectrum of paediatric neurotransmitter diseases and infantile parkinsonism. J Inherit Metab Dis 2009;32:321-32.7. Pons R, Ford B, Chiriboga CA, et al. Aromatic L-amino acid decarboxylase deficiency: clinical features, treatment,
and prognosis. Neurology 2004;62:1058-65.8. Swoboda KJ, Hyland K, Goldstein DS, et al. Clinical and therapeutic observations in aromatic L-amino acid decarboxylase deficiency. Neurology 1999;53:1205-11.9. Yeung WL, Wong VC, Chan KY, et al. Expanding phenotype and clinical analysis of tyrosine hydroxylase deficiency. J Child Neurol 2011;26:179-87. [Erratum, J Child Neurol 2012;27:829-31.]10. Kurian MA, Zhen J, Cheng SY, et al. Homozygous loss-of-function mutations in the gene encoding the dopamine trans-porter are associated with infantile par-
The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission.
Copyright © 2013 Massachusetts Medical Society. All rights reserved.

n engl j med 368;6 nejm.org february 7, 2013550
brief report
kinsonism-dystonia. J Clin Invest 2009; 119:1595-603.11. Burman J, Tran CH, Glatt C, Freimer NB, Edwards RH. The effect of rare hu-man sequence variants on the function of vesicular monoamine transporter 2. Pharmacogenetics 2004;14:587-94.12. Glatt CE, DeYoung JA, Delgado S, et al. Screening a large reference sample to identify very low frequency sequence vari-ants: comparisons between two genes. Nat Genet 2001;27:435-8.13. Iwasa H, Kurabayashi M, Nagai R, Nakamura Y, Tanaka T. Multiple single-nucleotide polymorphisms (SNPs) in the Japanese population in six candidate genes for long QT syndrome. J Hum Genet 2001;46:158-62.14. Erickson JD, Eiden LE. Functional identification and molecular cloning of a human brain vesicle monoamine trans-porter. J Neurochem 1993;61:2314-7.15. Partridge AW, Therien AG, Deber CM. Missense mutations in transmembrane domains of proteins: phenotypic propen-sity of polar residues for human disease. Proteins 2004;54:648-56.
16. Duerr JS, Frisby DL, Gaskin J, et al. The cat-1 gene of Caenorhabditis elegans encodes a vesicular monoamine transport-er required for specific monoamine-depen-dent behaviors. J Neurosci 1999;19:72-84.17. Eiden LE, Weihe E. VMAT2: a dynamic regulator of brain monoaminergic neuro-nal function interacting with drugs of abuse. Ann N Y Acad Sci 2011;1216:86-98.18. Dayhoff MO, Schwartz RM, Orcutt BC. A model of evolutionary change in proteins. In: Dayhoff MO, ed. Atlas of protein sequence and structure. Washing-ton, DC: National Biomedical Research Foundation, 1978:345-52.19. Henikoff S, Henikoff JG. Amino acid substitution matrices from protein blocks. Proc Natl Acad Sci U S A 1992;89:10915-9.20. Grantham R. Amino acid difference formula to help explain protein evolution. Science 1974;185:862-4.21. Wang YM, Gainetdinov RR, Fuma-galli F, et al. Knockout of the vesicular monoamine transporter 2 gene results in neonatal death and supersensitivity to cocaine and amphetamine. Neuron 1997; 19:1285-96.
22. Fon EA, Pothos EN, Sun BC, Killeen N, Sulzer D, Edwards RH. Vesicular transport regulates monoamine storage and release but is not essential for am-phetamine action. Neuron 1997;19:1271-83.23. Mooslehner KA, Chan PM, Xu W, et al. Mice with very low expression of the ve-sicular monoamine transporter 2 gene survive into adulthood: potential mouse model for parkinsonism. Mol Cell Biol 2001;21:5321-31.24. Fukui M, Rodriguiz RM, Zhou J, et al. Vmat2 heterozygous mutant mice display a depressive-like phenotype. J Neurosci 2007;27:10520-9.25. Jeon SM, Cheon SM, Bae HR, Kim JW, Kim SU. Selective susceptibility of human dopaminergic neural stem cells to dopa-mine-induced apoptosis. Exp Neurobiol 2010;19:155-64.26. Bainbridge MN, Wiszniewski W, Mur-dock DR, et al. Whole-genome sequenc-ing for optimized patient management. Sci Transl Med 2011;3(87):87re3.Copyright © 2013 Massachusetts Medical Society.
The New England Journal of Medicine Downloaded from nejm.org by KHALED ABDELNASER on February 9, 2013. For personal use only. No other uses without permission.
Copyright © 2013 Massachusetts Medical Society. All rights reserved.
Related Documents