Antimicrobial Peptides as Novel Alternatives to Antibiotics in Swine and Poultry Industries F.J. Ji 1,2 , H.S. Yang 1,3 *, Z.Y. Cui 4 and Y.L. Yin 1,3 * 1 Animal Nutrition and Human Health Laboratory, School of Life Sciences, Hunan Normal University, 410081Changsha, Hunan, China 2 Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences, 571737 Danzhou, Hainan, China 3 Key Laboratory for Agro-Ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, the Chinese Academy of Sciences, 410125 Changsha, Hunan, China 4 Guangdong Hinabiotech Co., Ltd. 511400 Guangzhou, Guangdong, China * Corresponding to H.S. Yang ([email protected]) or Y.L. Yin ([email protected]) There is a worldwide concern that bacterial resistance to antibiotics has endangered public health. Alternative antibacterial agents need to be sought to maintain animal health and human food safety. Antimicrobial peptides (AMPs) as a part of the nonspecific defence system, can be produced by many organisms, from bacteria to plants, insects, amphibians and mammals, as well as by chemical synthesis. The article investigates the structure and mechanism of action of AMPs, especially their effects on growth performance, nutrient digestibility, intestinal health and immune modulation, gut microbiota in non-ruminant nutrition. Overall, due to their broad-spectrum microbial activity against various fungi, bacteria and enveloped viruses, AMPs show considerable potential as feed additives replacing antibiotics applied in swine and poultry industries. However, research is still needed in this area, considering the pharmacokinetic and drug-resistance of AMPs were poorly understood. Keywords: antimicrobial peptides; alternatives to antibiotics; applications; swine; poultry 1. Background Antibiotics are typically used as a cost-effective tool for improving feed efficiency and for preventing diseases in animal feed industry[1]. However, their continuous use and misuse of antibiotics has led to the emergence of bacteria resistant, and consumers are becoming increasingly concerned about drug residues in meat products[2]. Moreover, with the emergence of the first plasmid-mediated polymyxin resistance, designated mcr-1, in Enterobacteria from animal and human isolates, the scientist provide evidence for the horizontal dissemination of mcr-1 from the veterinary sector to human beings[3]. Therefore, antimicrobial resistance has been recognized one of the most serious global threats to human health. To address antibacterial resistance, the government has started to ban the inclusion of conventional antibiotics in animal diets as a routine means of growth promotion. In addition, an intensive amount of research has been implemented on the development of alternatives to antibiotics to maintain animal performance and health in the past two decades. Among of these, the top ten approaches are antibodies, probiotics, bacteriophages (wild type and engineered respectively), vaccines, immune stimulation, and antimicrobial peptides, host defense peptide and antibiofilm peptides are also involved[4]. In this article, we investigate the categories and mechanism of action of AMPs, review their effects on growth performance, nutrient digestibility, intestinal health, immune status and gut microbiota. 2. Categories of AMPs AMPs encompass a great diversity of biology and molecules, based on their different sources, biological activities, amino acids make-up and structures. There are so many ways to classify AMPs to different group according to different definition. Usually, we refer to some database, such as an online antimicrobial peptide database(APD3, accessible at http://aps.unmc.edu/AP/[5]. The APD3 database currently lists over 2600 examples of AMPs from all kingdoms of life: bacteria, archaea, and eukaryotes (including plants, animals, fungi and protists). AMPs are potent, broad function which contains antibacterial, antiviral, anti-HIV, antifungal, antiparasitic and anticancer peptides. Currently, more than seventy-five percentage of AMPs with several different sequences have been isolated from a wide range of animals. Newly annotated are AMPs with antibiofilm, antimalarial, anti-protist, insecticidal, spermicidal, chemotactic, wound healing, antioxidant and protease inhibiting properties[6]. In addition, amino acid composition of natural AMPs is very important for peptide classification and prediction, especially design, recombinant. According to the three-dimensional structures of AMPs, they can be classified into four major classes: α-helix, β- sheet, extended and loop peptides (figure1). Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.) 19 _____________________________________________________________________________

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Antimicrobial Peptides as Novel Alternatives to Antibiotics in Swine and Poultry Industries
F.J. Ji1,2, H.S. Yang1,3*, Z.Y. Cui4 and Y.L. Yin1,3* 1 Animal Nutrition and Human Health Laboratory, School of Life Sciences, Hunan Normal University, 410081Changsha,
Hunan, China 2 Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences, 571737 Danzhou,
Hainan, China 3 Key Laboratory for Agro-Ecological Processes in Subtropical Region, Institute of Subtropical Agriculture, the Chinese
Academy of Sciences, 410125 Changsha, Hunan, China
4 Guangdong Hinabiotech Co., Ltd. 511400 Guangzhou, Guangdong, China
*Corresponding to H.S. Yang ([email protected]) or Y.L. Yin ([email protected])
There is a worldwide concern that bacterial resistance to antibiotics has endangered public health. Alternative antibacterial agents need to be sought to maintain animal health and human food safety. Antimicrobial peptides (AMPs) as a part of the nonspecific defence system, can be produced by many organisms, from bacteria to plants, insects, amphibians and mammals, as well as by chemical synthesis. The article investigates the structure and mechanism of action of AMPs, especially their effects on growth performance, nutrient digestibility, intestinal health and immune modulation, gut microbiota in non-ruminant nutrition. Overall, due to their broad-spectrum microbial activity against various fungi, bacteria and enveloped viruses, AMPs show considerable potential as feed additives replacing antibiotics applied in swine and poultry industries. However, research is still needed in this area, considering the pharmacokinetic and drug-resistance of AMPs were poorly understood.
Keywords: antimicrobial peptides; alternatives to antibiotics; applications; swine; poultry
1. Background
Antibiotics are typically used as a cost-effective tool for improving feed efficiency and for preventing diseases in animal feed industry[1]. However, their continuous use and misuse of antibiotics has led to the emergence of bacteria resistant, and consumers are becoming increasingly concerned about drug residues in meat products[2]. Moreover, with the emergence of the first plasmid-mediated polymyxin resistance, designated mcr-1, in Enterobacteria from animal and human isolates, the scientist provide evidence for the horizontal dissemination of mcr-1 from the veterinary sector to human beings[3]. Therefore, antimicrobial resistance has been recognized one of the most serious global threats to human health. To address antibacterial resistance, the government has started to ban the inclusion of conventional antibiotics in animal diets as a routine means of growth promotion. In addition, an intensive amount of research has been implemented on the development of alternatives to antibiotics to maintain animal performance and health in the past two decades. Among of these, the top ten approaches are antibodies, probiotics, bacteriophages (wild type and engineered respectively), vaccines, immune stimulation, and antimicrobial peptides, host defense peptide and antibiofilm peptides are also involved[4]. In this article, we investigate the categories and mechanism of action of AMPs, review their effects on growth performance, nutrient digestibility, intestinal health, immune status and gut microbiota.
2. Categories of AMPs
AMPs encompass a great diversity of biology and molecules, based on their different sources, biological activities, amino acids make-up and structures. There are so many ways to classify AMPs to different group according to different definition. Usually, we refer to some database, such as an online antimicrobial peptide database(APD3, accessible at http://aps.unmc.edu/AP/[5]. The APD3 database currently lists over 2600 examples of AMPs from all kingdoms of life: bacteria, archaea, and eukaryotes (including plants, animals, fungi and protists). AMPs are potent, broad function which contains antibacterial, antiviral, anti-HIV, antifungal, antiparasitic and anticancer peptides. Currently, more than seventy-five percentage of AMPs with several different sequences have been isolated from a wide range of animals. Newly annotated are AMPs with antibiofilm, antimalarial, anti-protist, insecticidal, spermicidal, chemotactic, wound healing, antioxidant and protease inhibiting properties[6]. In addition, amino acid composition of natural AMPs is very important for peptide classification and prediction, especially design, recombinant. According to the three-dimensional structures of AMPs, they can be classified into four major classes: α-helix, β-sheet, extended and loop peptides (figure1).
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
19
_____________________________________________________________________________

Figure 1. Protein models representing the structural differences of the four classes of antimicrobial peptides. Antimicrobial peptides can be grouped into four major classes based on their secondary structures, including the (A) alfa-helical peptides, (B) beta-sheets peptides, (C) peptides that adopt unconventional structures, such as extended helices, and (D) peptides that assemble into loops. All structures were obtained freely from the RCSB Protein Data Bank (PDB) (http://www.pdb.org/) [7]and have been referenced according to their Digital Object Identifier [8]. Additional information for each AMP may be obtained by consulting the RCSB PDB and cross-referencing Peters et al[9].
3. Differ AMPs from peptide-based antibiotics
Similar to traditional peptide-based antibiotics, antimicrobial peptides have been described as evolutionarily ancient weapons against microbial infections, with a variable composition of amino acids, target on membrane or intracellular. Actually, in some scientist mind, those natural peptides synthesized gene coded are true AMPs. Compared with peptide-based antibiotics produced by microorganism, AMPs mainly differ with them at synthesis process and amino acid make-up although they are peptide-based small molecules. Firstly, those peptide-based antibiotics are produced by non-ribosomal peptide synthesis (NRPS). During this biology process, multiple enzymes encoded by the expression of large arrays of genes are involved. The enzymes work in succession to catalyse the sequence of chemical reactions required to synthetize the antibiotics [10, 11]. For example, β-lactams such as penicillin, cyclic peptide antibiotics like the polymyxins and bacitracin, glycopeptides like vancomycin [12], are grouped into peptide-based antibiotics. In contrast to NRPS-antibiotics, AMPs including those made in human cells, are produced by the normal process of ribosomal translation on an mRNA template. The primary product is usually a preprotein that is then processed to the final length of the active AMPs. Secondly, AMPs and NRPS-antibiotics have different consequences for the compositions of the final products. Ribosomal produced AMPs contain only the normal complement of amino acids found in proteins, sometimes with post-translational modifications. NRPS-antibiotics, in contrast, are unconstrained by the limitations of ribosomal translation, and usually contain mixture of normal amino acids together with non-canonical amino acids not found in any proteins[13].
4. Mechanisms of AMPs action
Considering the special proprieties of AMPs, like variable composition of amino acids, net charge, spatial conformation and structure, amphipathic modules and fluidity of membrane, AMPs were believed to possess a special mechanism of action. The mechanism of the antimicrobial activities of AMPs has been studied for some selected peptides. Based on the results, there have two main kinds of action mode of AMPs namely membrane interaction specificity and intracellular targets.
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
20
_____________________________________________________________________________

4.1 Membrane specific targets
The bactericidal effect of AMPs is widely believed to be due to the formation of pores in the bacterial cytoplasmic membrane, resulting in a loss of control over ion flows across the membrane and cell death. To supporting this, several models for AMP-directed membrane pore formation have been suggested: (1) the barrel-stave model;(2) the carpet model; (3) the toroidal pore model[14]. Recently, increasing evidence shows that the interaction between different AMPs and bacterial membranes may involve much more specific interactions than the generic models of pore formation[15]. For example, phosphatidylethanolamine (PE), present on the surface of bacterial membranes, acts as a high affinity lipid receptor for several AMPs [16]. Lipopolysaccharides(LPS), lipoteichoic acid (LTA) and the peptidoglycan precursor lipid II were also specific targeted by AMPs during the course of interaction with bacterial membranes [17]. Precursor lipid II seems to be a favorite target. It has been demonstrated that plectasin, a fungal defensin, acts by directly binding the bacterial cell-wall precursor Lipid II[18]. Another clue to the specificity of killing bacteria by membrane disruption is that there are significant differences in the lipid composition of bacterial (like gram-negative bacteria, gram-positive bacteria) and eukaryotic cell (e.g. human cell )membranes[19, 20].
4.2 Intracellular targets
Recent research also shows that at least part of antimicrobial activity of AMPs might be exerted in the cytoplasm[21]. Indolicidin, a bovine cathelicidin, has been shown not to lyse bacterial cells but to inhibit DNA synthesis[22] and shown in vitro to interact with duplex DNA[23]. Analogs of buforin antimicrobial peptides were also shown to exert their bactericidal activity on E. Coli by binding to DNA and RNA after penetrating the cell membrane[24]. Objectively, we only discuss two main kind of mechanisms of AMPs for your reference since there are so many different AMPs in the world.
5. The application of AMPs in swine and poultry nutrition
With the increasing concern about microbial resistance to conventional antibiotics, AMPs have been more and more attracting in swine and poultry industries. In the past decade, many AMPs, including antimicrobial peptide cecropin, plectasin, and sublancin, cathelicidin have been extensively evaluated effects on growth performance, nutrient digestibility, immune status and intestinal health as well as fecal microbiota, especially under some special stress conditions. The studies suggesting that AMPs is could be considered as novel alternatives to antibiotic growth promoters. The detailed results are summarized in table1,2,3 respectively.
5.1 Promote growth performance
Previous studies have demonstrated the benefits of AMPs on the growth performance of swine and poultry (Table 1). For piglets, those studies displayed AMPs had better performance and lower diarrheal rate than both basal diet group[25-27] and challenged group[28], although the performance parameters showed a little lower less antibiotic group[27, 28]. Furthermore, some trial showed that the pigs fed with AMPs could maintain the similar performance and fecal score to antibiotic group [29-34], even challenged by E. Coli [35, 36]. In addition, AMPs sometimes demonstrated better performance in contrast to antibiotic group by increasing body weight and decreasing diarrheal rate [37, 38]. According to the summary, all of studies were focused on weanling pigs, that is in accordance with the animal industry. Actually, most majority of antibiotics were used during the period from post-weanling to 30 kg body weight. For broilers, most studies were focused on overall of the production cycle. That was because a number of antibiotic used in the birds was only banned one week before arriving marketing body weight considering its very short feeding time and high incidence of disease. Moreover, the studies suggesting that supplementation with AMPs showed better performance than basal diet group[39, 40], and could resist the adverse effects induced by C. Perfringens or heat stress and maintain similar growth performance to control group [41, 42]. Similarly, either under normal conditions[43, 44], or challenge model[45], the diet added with AMPs had the similar performance to antibiotic diet.
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
21
_____________________________________________________________________________
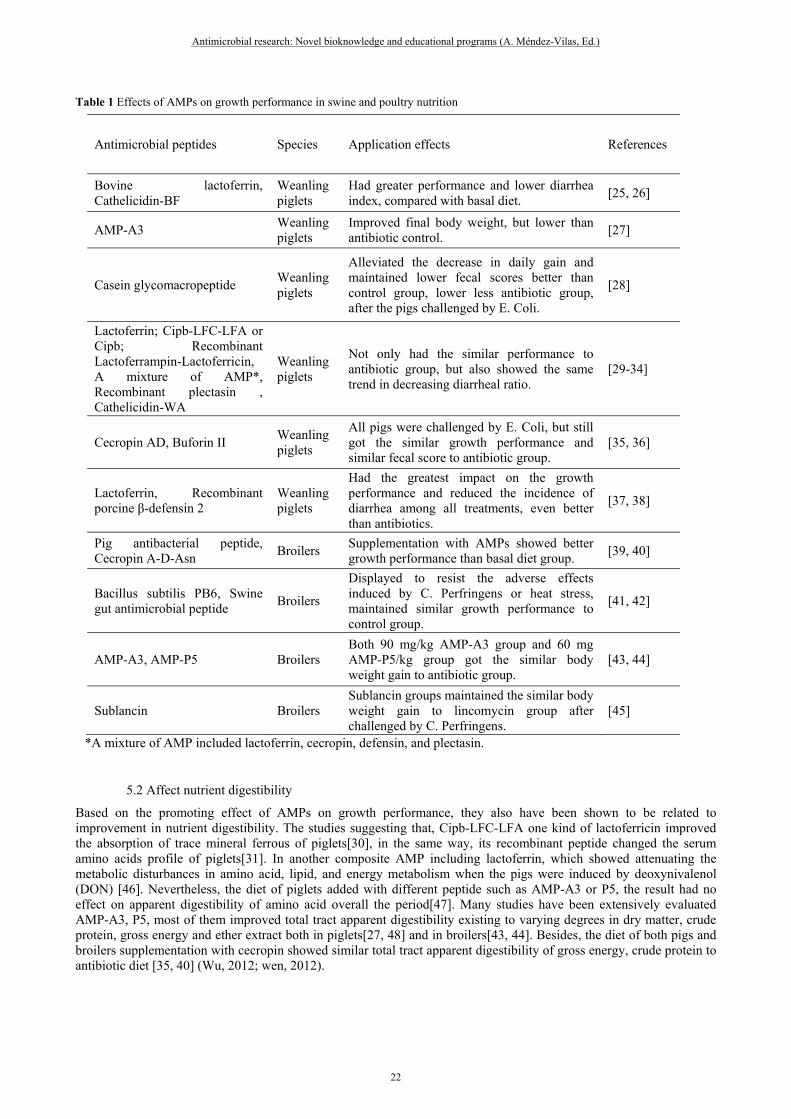
Table 1 Effects of AMPs on growth performance in swine and poultry nutrition
Antimicrobial peptides Species Application effects References
Bovine lactoferrin, Cathelicidin-BF
Weanling piglets
Had greater performance and lower diarrhea index, compared with basal diet.
[25, 26]
AMP-A3 Weanling piglets
Improved final body weight, but lower than antibiotic control.
[27]
Casein glycomacropeptide Weanling piglets
Alleviated the decrease in daily gain and maintained lower fecal scores better than control group, lower less antibiotic group, after the pigs challenged by E. Coli.
[28]
Lactoferrin; Cipb-LFC-LFA or Cipb; Recombinant Lactoferrampin-Lactoferricin, A mixture of AMP*, Recombinant plectasin , Cathelicidin-WA
Weanling piglets
Not only had the similar performance to antibiotic group, but also showed the same trend in decreasing diarrheal ratio.
[29-34]
Cecropin AD, Buforin II Weanling piglets
All pigs were challenged by E. Coli, but still got the similar growth performance and similar fecal score to antibiotic group.
[35, 36]
Lactoferrin, Recombinant porcine β-defensin 2
Weanling piglets
Had the greatest impact on the growth performance and reduced the incidence of diarrhea among all treatments, even better than antibiotics.
[37, 38]
Pig antibacterial peptide, Cecropin A-D-Asn
Broilers Supplementation with AMPs showed better growth performance than basal diet group.
[39, 40]
Bacillus subtilis PB6, Swine gut antimicrobial peptide
Broilers
Displayed to resist the adverse effects induced by C. Perfringens or heat stress, maintained similar growth performance to control group.
[41, 42]
AMP-A3, AMP-P5 Broilers Both 90 mg/kg AMP-A3 group and 60 mg AMP-P5/kg group got the similar body weight gain to antibiotic group.
[43, 44]
Sublancin Broilers Sublancin groups maintained the similar body weight gain to lincomycin group after challenged by C. Perfringens.
[45]
*A mixture of AMP included lactoferrin, cecropin, defensin, and plectasin.
5.2 Affect nutrient digestibility
Based on the promoting effect of AMPs on growth performance, they also have been shown to be related to improvement in nutrient digestibility. The studies suggesting that, Cipb-LFC-LFA one kind of lactoferricin improved the absorption of trace mineral ferrous of piglets[30], in the same way, its recombinant peptide changed the serum amino acids profile of piglets[31]. In another composite AMP including lactoferrin, which showed attenuating the metabolic disturbances in amino acid, lipid, and energy metabolism when the pigs were induced by deoxynivalenol (DON) [46]. Nevertheless, the diet of piglets added with different peptide such as AMP-A3 or P5, the result had no effect on apparent digestibility of amino acid overall the period[47]. Many studies have been extensively evaluated AMP-A3, P5, most of them improved total tract apparent digestibility existing to varying degrees in dry matter, crude protein, gross energy and ether extract both in piglets[27, 48] and in broilers[43, 44]. Besides, the diet of both pigs and broilers supplementation with cecropin showed similar total tract apparent digestibility of gross energy, crude protein to antibiotic diet [35, 40] (Wu, 2012; wen, 2012).
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
22
_____________________________________________________________________________
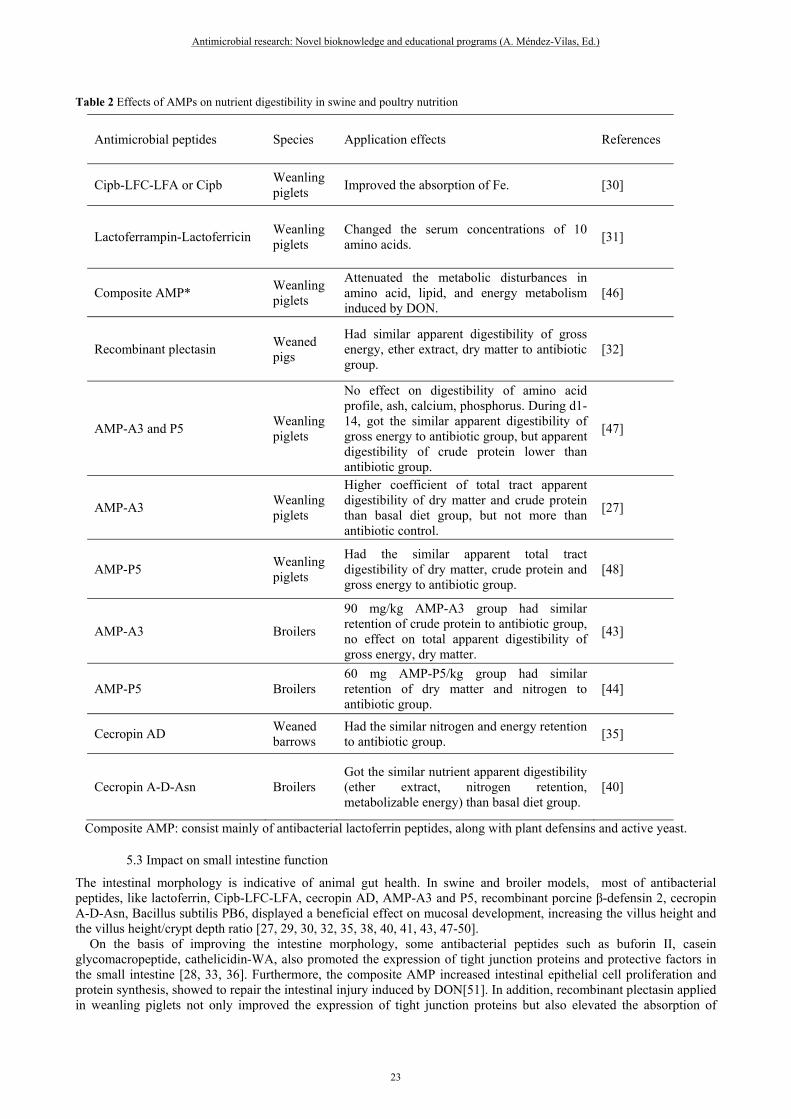
Table 2 Effects of AMPs on nutrient digestibility in swine and poultry nutrition
Antimicrobial peptides Species Application effects References
Cipb-LFC-LFA or Cipb Weanling piglets
Improved the absorption of Fe. [30]
Lactoferrampin-Lactoferricin Weanling piglets
Changed the serum concentrations of 10 amino acids.
[31]
Composite AMP* Weanling piglets
Attenuated the metabolic disturbances in amino acid, lipid, and energy metabolism induced by DON.
[46]
Recombinant plectasin Weaned pigs
Had similar apparent digestibility of gross energy, ether extract, dry matter to antibiotic group.
[32]
AMP-A3 and P5 Weanling piglets
No effect on digestibility of amino acid profile, ash, calcium, phosphorus. During d1-14, got the similar apparent digestibility of gross energy to antibiotic group, but apparent digestibility of crude protein lower than antibiotic group.
[47]
AMP-A3 Weanling piglets
Higher coefficient of total tract apparent digestibility of dry matter and crude protein than basal diet group, but not more than antibiotic control.
[27]
AMP-P5 Weanling piglets
Had the similar apparent total tract digestibility of dry matter, crude protein and gross energy to antibiotic group.
[48]
AMP-A3 Broilers
90 mg/kg AMP-A3 group had similar retention of crude protein to antibiotic group, no effect on total apparent digestibility of gross energy, dry matter.
[43]
AMP-P5 Broilers 60 mg AMP-P5/kg group had similar retention of dry matter and nitrogen to antibiotic group.
[44]
Cecropin AD Weaned barrows
Had the similar nitrogen and energy retention to antibiotic group.
[35]
Cecropin A-D-Asn Broilers Got the similar nutrient apparent digestibility (ether extract, nitrogen retention, metabolizable energy) than basal diet group.
[40]
Composite AMP: consist mainly of antibacterial lactoferrin peptides, along with plant defensins and active yeast.
5.3 Impact on small intestine function
The intestinal morphology is indicative of animal gut health. In swine and broiler models, most of antibacterial peptides, like lactoferrin, Cipb-LFC-LFA, cecropin AD, AMP-A3 and P5, recombinant porcine β-defensin 2, cecropin A-D-Asn, Bacillus subtilis PB6, displayed a beneficial effect on mucosal development, increasing the villus height and the villus height/crypt depth ratio [27, 29, 30, 32, 35, 38, 40, 41, 43, 47-50]. On the basis of improving the intestine morphology, some antibacterial peptides such as buforin II, casein glycomacropeptide, cathelicidin-WA, also promoted the expression of tight junction proteins and protective factors in the small intestine [28, 33, 36]. Furthermore, the composite AMP increased intestinal epithelial cell proliferation and protein synthesis, showed to repair the intestinal injury induced by DON[51]. In addition, recombinant plectasin applied in weanling piglets not only improved the expression of tight junction proteins but also elevated the absorption of
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
23
_____________________________________________________________________________

xylose, plasma disaccharides activity in the small intestine. Whilst, another trials suggesting that antimicrobial peptides such as pig AMP and swine gut AMP increased the activity of intestinal alkaline phosphatase in broilers [39, 52]. However, for diamine oxide, d-lactate and entothelin-1, the indicators of intestinal permeability, dietary supplementation with recombinant Lactoferrampin-Lactoferricin showed no significant effects among all treatments[31].
5.4 Regulate immunity
It is reported that AMPs are effector molecules of innate and adaptive immunity with modulation of pro- and anti-inflammatory responses, chemotactic activity, and direct effects on adaptive immunity [53-55]. Increasing evidence indicated that the addition of AMPs strengthened host defenses by increasing lymphocyte proliferation, the level of IgG, IgA, IgM and cytokines in serum, the expression of secretory IgA [31, 37, 39, 49, 56]. In pigs, antibacterial peptide cathelicidin-BF inhibited the expression of the inflammatory cytokines but promoted the expression of the anti-inflammatory cytokine in the intestine [26]. Recently, there has been increase effort to investigate the interaction of cationic peptides with host cells as immunomodulators in mammals [57-59]. Ren et al, reported that a composite AMP including swine defensin and fly antimicrobial peptide increased the T cell population and proliferation function and decreased percentages of apoptotic cells in broilers[60]. In vitro, chicken macrophage cell line HD11 and chicken primary monocytes were stimulated with various concentrations of the chicken cathelicidin-2 (CATH-2) in a range of 10-40 umol, the data suggested that CATH-2 increased the transcription of chemokines, but no effect on pro-inflammatory cytokine IL-1β[61]. Similarly, using various concentrations of chicken NK-lysine-derived peptide (CNK-2) in a range of 10-300 ug/ml in the same cell lines model, the result showed that 300 ug/ml CNK-2 had cytoxic effect on HD11 cells but no effect on monocytes and suppressed the expression of pro-inflammatory cytokine in HD11 cells and in primary macrophage cells[62]. These results indicated that CATH-2 and CNK-2 are potential novel immunomodulating agent rather than antimicrobial agent.
5.5 Modulate gut microbiota
The beneficial effects of AMPs on the host animal have been demonstrated by previous studies (table 3). The results indicated that AMPs modulated the microbiota ecology by suppressing harmful microorganisms like E. Coli and Clostridium and promoting proliferation of beneficial microorganisms like Lactobacillus and Bifidobacterium [29, 30, 32]. Moreover, microbiota concentration in pigs was affected in a dose-dependent manner. It was reported that, with the increasing level of antibacterial peptide A3, the populations of total anaerobic bacteria, coliforms, Clostridium spp. in feces, ileum and cecum were linearly reduced[27]. The dose-dependent effects of antibacterial peptide A3 and P5 on gut microbiota have also been observed in broilers [43, 44]. Besides, we also pay attention to the different effects between AMPs and the conventional antibiotics on intestinal microbiota, and keep concerned with short chain volatile fatty acids what were produced by gut microbiota during the course of energy metabolism. In diarrhea piglets, the basal diet was designed to add with antibacterial peptide cathelicidin-WA (CWA) and enrofloxacin (Enro), respectively. Compared with basal diet, both CWA and Enro diet decreased the ratio of Escherichia coli to total bacteria and increased the ratio of Lactobacillus to Escherichia coli in feces; CWA increased acetate, propionate and butyrate concentrations in feces. By contrast, the ratio of Lactobacillus to total bacteria in feces was deceased by Enro but increased by CWA compared with the control. Furthermore, cathelicidin-WA increased acetate, propionate and butyrate concentrations, but Enro decreased butyrate concentration compared with the control. The result indicated that AMPs had more positive effecting on regulation of gut microflora in contrast to conventional antibiotics[33]. In broilers, antimicrobial peptide sublancin significantly decreased the cecum Clostridium perfringens in challenged broilers. In contrast to lincomycin group, sublancin treatment had the highest Lactobacilli counts which may be an advantage for AMPs compared to traditional antibiotics in regulation of gut microflora[45].
6. Conclusion
The use of AMPs in animals has provided several benefits, like growth performance, nutrient digestibility, gut health, intestinal microflora and immunomodulation, to the swine and poultry industry. However, we noted that AMPs are not omnipotent. It is inevitable that innate resistance and acquired resistance occur in some AMPs species since they encounter bacteria frequently in their natural environments and promotor use. For instance, Staphylococcus aureus isolated from clinical sample shows reducing susceptibility to LL-37 and human defensins [64]. Recently, the emergence of mcr-1 prompts us to note that acquired resistance to colistin from swine to human [3]. Furthermore, the actual mode of AMPs metabolism in vivo remains an area where there is still a poor level of understanding. Nowadays our further research will advance knowledge within these fields and highlight the potential of AMPs as alternatives to conventional antibiotics. Eventually, AMPs may become useful growth promotor and therapeutic agents, since they have shown not only antibacterial, but also immunomodulatory.
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
24
_____________________________________________________________________________
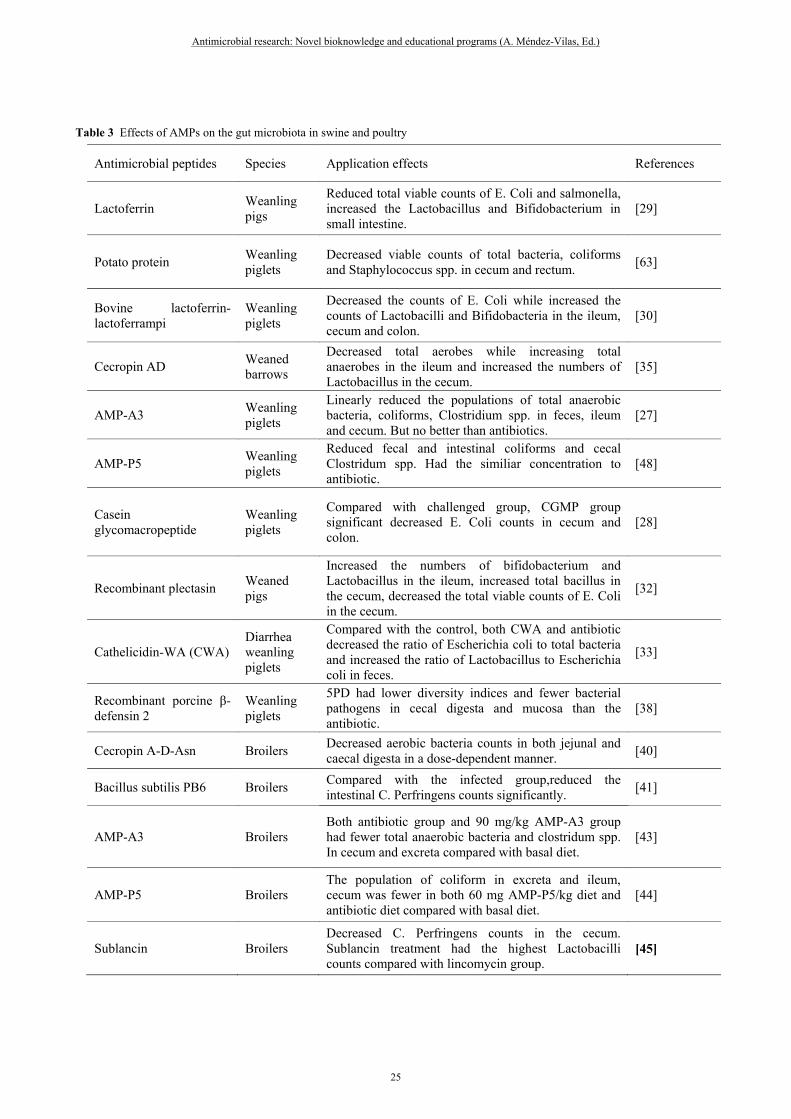
Table 3 Effects of AMPs on the gut microbiota in swine and poultry
Antimicrobial peptides Species Application effects References
Lactoferrin Weanling pigs
Reduced total viable counts of E. Coli and salmonella, increased the Lactobacillus and Bifidobacterium in small intestine.
[29]
Potato protein Weanling piglets
Decreased viable counts of total bacteria, coliforms and Staphylococcus spp. in cecum and rectum.
[63]
Bovine lactoferrin-lactoferrampi
Weanling piglets
Decreased the counts of E. Coli while increased the counts of Lactobacilli and Bifidobacteria in the ileum, cecum and colon.
[30]
Cecropin AD Weaned barrows
Decreased total aerobes while increasing total anaerobes in the ileum and increased the numbers of Lactobacillus in the cecum.
[35]
AMP-A3 Weanling piglets
Linearly reduced the populations of total anaerobic bacteria, coliforms, Clostridium spp. in feces, ileum and cecum. But no better than antibiotics.
[27]
AMP-P5 Weanling piglets
Reduced fecal and intestinal coliforms and cecal Clostridum spp. Had the similiar concentration to antibiotic.
[48]
Casein glycomacropeptide
Weanling piglets
Compared with challenged group, CGMP group significant decreased E. Coli counts in cecum and colon.
[28]
Recombinant plectasin Weaned pigs
Increased the numbers of bifidobacterium and Lactobacillus in the ileum, increased total bacillus in the cecum, decreased the total viable counts of E. Coli in the cecum.
[32]
Cathelicidin-WA (CWA) Diarrhea weanling piglets
Compared with the control, both CWA and antibiotic decreased the ratio of Escherichia coli to total bacteria and increased the ratio of Lactobacillus to Escherichia coli in feces.
[33]
Recombinant porcine β-defensin 2
Weanling piglets
5PD had lower diversity indices and fewer bacterial pathogens in cecal digesta and mucosa than the antibiotic.
[38]
Cecropin A-D-Asn Broilers Decreased aerobic bacteria counts in both jejunal and caecal digesta in a dose-dependent manner.
[40]
Bacillus subtilis PB6 Broilers Compared with the infected group,reduced the intestinal C. Perfringens counts significantly.
[41]
AMP-A3 Broilers Both antibiotic group and 90 mg/kg AMP-A3 group had fewer total anaerobic bacteria and clostridum spp. In cecum and excreta compared with basal diet.
[43]
AMP-P5 Broilers The population of coliform in excreta and ileum, cecum was fewer in both 60 mg AMP-P5/kg diet and antibiotic diet compared with basal diet.
[44]
Sublancin Broilers Decreased C. Perfringens counts in the cecum. Sublancin treatment had the highest Lactobacilli counts compared with lincomycin group.
[45]
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
25
_____________________________________________________________________________

Acknowledgements We thank Andy Hu for reading and commenting on this manuscript. This work was supported by National Natural Science Foundation of China (No. 31402089, 31330075), Key Programs of Frontier Scientific Research of the Chinese Academy of Sciences (QYZDY-SSW- SMC008).
Author Contributions Feng-jie Ji carried out the literature study and draft the manuscript. Huan-sheng Yang, Zhi-ying Cui and Yu-long Yin critically evaluated the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest The authors declare no conflict of interest.
References [1] Allen HK, Torey L, Bayles DO, Samuel H, Levine UY, David A, et al. Antibiotics in feed induce prophages in swine fecal
microbiomes. mBio by American Society for Microbiology. 2011;2:1867-77. [2] Thacker PA. Alternatives to antibiotics as growth promoters for use in swine production: A review. Journal of animal science and
biotechnology. 2014;4:35. [3] Liu YY, Wang Y, Walsh TR, Yi LX, Zhang R, Spencer J, et al. Emergence of plasmid-mediated colistin resistance mechanism
mcr-1 in animals and human beings in china: A microbiological and molecular biological study. Lancet Infectious Diseases. 2016;16:161.
[4] Czaplewski L, Bax R, Clokie M, Dawson M, Fairhead H, Fischetti VA, et al. Alternatives to antibiotics—a pipeline portfolio review. Lancet Infectious Diseases. 2016;16:239-51.
[5] Wang G. The antimicrobial peptide database. http://aps.unmc.edu/AP/2017. [6] Wang G, Li X, Wang Z. Apd3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Research.
2015;44:D1087–93. [7] Rcsb protein data bank (pdb). [8] Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H, et al. The protein data bank/ nucleic acids research. Nucleic
Acids Research. 2000;17:3588. [9] Peters BM, Shirtliff ME, Jabrarizk MA. Antimicrobial peptides: Primeval molecules or future drugs? Plos Pathogens.
2010;6:e1001067. [10] Baltz RH. Molecular engineering approaches to peptide, polyketide and other antibiotics. Nature Biotechnology. 2006;24:1533. [11] Hughes D. Exploiting genomics, genetics and chemistry to combat antibiotic resistance. Nature Reviews Genetics. 2003;4:432-
41. [12] Yim G, Thaker MN, Koteva K, Wright G. Glycopeptide antibiotic biosynthesis. Journal of Antibiotics. 2014;67:31. [13] Walsh CT, O'Brien RV, Khosla C. Cheminform abstract: Nonproteinogenic amino acid building blocks for nonribosomal
peptide and hybrid polyketide scaffolds. Angewandte Chemie International Edition. 2013;52:7098. [14] Melo MN, Ferre R, Castanho MA. Antimicrobial peptides: Linking partition, activity and high membrane-bound concentrations.
Nature Reviews Microbiology. 2009;7:245-50. [15] Andersson DI, Hughes D, Kubicek-Sutherland JZ. Mechanisms and consequences of bacterial resistance to antimicrobial
peptides. Drug Resistance Updates. 2016;26:43-57. [16] Phoenix DA, Harris F, Mura M, Dennison SR. The increasing role of phosphatidylethanolamine as a lipid receptor in the action
of host defence peptides. Progress in Lipid Research. 2015;59:26-37. [17] Schmitt P, Rosa RD, Destoumieuxgarzón D. An intimate link between antimicrobial peptide sequence diversity and binding to
essential components of bacterial membranes. Biochimica Et Biophysica Acta. 2015;1858:958-70. [18] Schneider T, Kruse T, Wimmer R, Wiedemann I, Sass V, Pag U, et al. Plectasin, a fungal defensin, targets the bacterial cell wall
precursor lipid ii. Science. 2010;328:1168-72. [19] Teixeira V, Feio MJ, Bastos M. Role of lipids in the interaction of antimicrobial peptides with membranes. Progress in Lipid
Research. 2012;51:149. [20] Sohlenkamp C, Geiger O. Bacterial membrane lipids: Diversity in structures and pathways. Fems Microbiology Reviews.
2015;40. [21] Chileveru HR, Lim SA, Chairatana P, Wommack AJ, Chiang IL, Nolan EM. Visualizing attack of escherichia coli by the
antimicrobial peptide human defensin 5. Biochemistry. 2015;54:1767. [22] Subbalakshmi C, Sitaram N. Mechanism of antimicrobial action of indolicidin. Fems Microbiology Letters. 1998;160:91-6. [23] Ghosh A, Kar RK, Jana J, Saha A, Jana B, Krishnamoorthy J, et al. Indolicidin targets duplex DNA: Structural and mechanistic
insight through a combination of spectroscopy and microscopy. Chemmedchem. 2014;9:2052. [24] Hao G, Shi YH, Tang YL, Le GW. The intracellular mechanism of action on escherichia coli of bf2-a/c, two analogues of the
antimicrobial peptide buforin 2. Journal of Microbiology. 2013;51:200-6. [25] Wang YZ, Xu CL, An ZH, Liu JX, Feng J. Effect of dietary bovine lactoferrin on performance and antioxidant status of piglets.
Animal Feed Science & Technology. 2008;140:326-36. [26] Yi H, Yu C, Zhang H, Song D, Jiang D, Du H, et al. Cathelicidin-bf suppresses intestinal inflammation by inhibiting the nuclear
factor-κb signaling pathway and enhancing the phagocytosis of immune cells via stat-1 in weanling piglets. International Immunopharmacology. 2015;28:61.
[27] Yoon JH, Ingale SL, Kim JS, Kim KH, Lee SH, Park YK, et al. Effects of dietary supplementation of antimicrobial peptide-a3 on growth performance, nutrient digestibility, intestinal and fecal microflora and intestinal morphology in weanling pigs. Animal Feed Science and Technology. 2012;177:98-107.
[28] Rong Y, Lu Z, Zhang H, Zhang L, Song D, Wang Y. Effects of casein glycomacropeptide supplementation on growth performance, intestinal morphology, intestinal barrier permeability and inflammatory responses in escherichia coli k88 challenged piglets. Animal Nutrition. 2015;1:54-9.
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
26
_____________________________________________________________________________

[29] Wang YZ, Shan TZ, Xu ZR, Feng J, Wang ZQ. Effects of the lactoferrin (lf) on the growth performance, intestinal microflora and morphology of weanling pigs. Animal Feed Science & Technology. 2007;135:263-72.
[30] Tang ZR, Yin YL, Zhang YM, Huang RL, Sun ZH, Li TJ, et al. Effects of dietary supplementation with an expressed fusion peptide bovine lactoferricin-lactoferrampin on performance, immune function and intestinal mucosal morphology in piglets weaned at age 21 d. British Journal of Nutrition. 2009;101:998-1005.
[31] Tang X, Fatufe AA, Yin Y, Tang Z, Wang S, Liu Z, et al. Dietary supplementation with recombinant lactoferrampin-lactoferricin improves growth performance and affects serum parameters in piglets. Journal of Animal & Veterinary Advances. 2012;11:2548-55.
[32] Wan J, Li Y, Chen D, Yu B, Chen G, Zheng P, et al. Recombinant plectasin elicits similar improvements in the performance and intestinal mucosa growth and activity in weaned pigs as an antibiotic. Animal Feed Science & Technology. 2016;211:216-26.
[33] Yi H, Zhang L, Gan Z, Xiong H, Yu C, Du H, et al. High therapeutic efficacy of cathelicidin-wa against postweaning diarrhea via inhibiting inflammation and enhancing epithelial barrier in the intestine. Scientific Reports. 2016;6:25679.
[34] Xiong X, Yang HS, Li L, Wang YF, Huang RL, Li FN, et al. Effects of antimicrobial peptides in nursery diets on growth performance of pigs reared on five different farms. Livestock Science. 2014;167:206-10.
[35] Wu S, Zhang F, Huang Z, Liu H, Xie C, Zhang J, et al. Effects of the antimicrobial peptide cecropin ad on performance and intestinal health in weaned piglets challenged with escherichia coli. Peptides. 2012;35:225-30.
[36] Tang ZR, Deng H, Zhang XL, Zen Y, Xiao DF, Sun WZ, et al. Effects of orally administering the antimicrobial peptide buforin ii on small intestinal mucosal membrane integrity, the expression of tight junction proteins and protective factors in weaned piglets challenged by enterotoxigenic escherichia coli. Animal Feed Science and Technology. 2013;186:177-85.
[37] Shan T, Wang Y, Wang Y, Liu J, Xu Z. Effect of dietary lactoferrin on the immune functions and serum iron level of weanling piglets. Journal of Animal Science. 2007;85:2140.
[38] Peng Z, Wang A, Xie L, Song W, Wang J, Yin Z, et al. Use of recombinant porcine β-defensin 2 as a medicated feed additive for weaned piglets. Scientific Reports. 2016;6:26790.
[39] Bao H, She R, Liu T, Zhang Y, Peng KS, Luo D, et al. Effects of pig antibacterial peptides on growth performance and intestine mucosal immune of broiler chickens. Poultry Science. 2009;88:291-7.
[40] Wen LF, He JG. Dose-response effects of an antimicrobial peptide, a cecropin hybrid, on growth performance, nutrient utilisation, bacterial counts in the digesta and intestinal morphology in broilers. British Journal of Nutrition. 2012;108:1756.
[41] Jayaraman S, Thangavel G, Kurian H, Mani R, Mukkalil R, Chirakkal H. Bacillus subtilis pb6 improves intestinal health of broiler chickens challenged with clostridium perfringens-induced necrotic enteritis. Poultry Science. 2013;92:370-4.
[42] Hu F, Gao X, She R, Chen J, Mao J, Xiao P, et al. Effects of antimicrobial peptides on growth performance and small intestinal function in broilers under chronic heat stress. Poultry Science. 2016;0:1-9.
[43] Choi SC, Ingale SL, Kim JS, Park YK, Kwon IK, Chae BJ. An antimicrobial peptide-a3: Effects on growth performance, nutrient retention, intestinal and faecal microflora and intestinal morphology of broilers. British Poultry Science. 2013;54:738-46.
[44] Choi SC, Ingale SL, Kim JS, Park YK, Kwon IK, Chae BJ. Effects of dietary supplementation with an antimicrobial peptide-p5 on growth performance, nutrient retention, excreta and intestinal microflora and intestinal morphology of broilers. Animal Feed Science & Technology. 2013;185:78-84.
[45] Wang S, Zeng XF, Wang QW, Zhu JL, Peng Q, Hou CL, et al. The antimicrobial peptide sublancin ameliorates necrotic enteritis induced by in broilers. Journal of Animal Science. 2015;93:4750-60.
[46] Xiao H, Wu MM, Shao FY, Tan BE, Li TJ, Ren WK, et al. Metabolic profiles in the response to supplementation with composite antimicrobial peptides in piglets challenged with deoxynivalenol. Journal of Animal Science. 2015;93:1114.
[47] Yoon JH, Ingale SL, Kim JS, Kim KH, Lee SH, Park YK, et al. Effects of dietary supplementation of synthetic antimicrobial peptide-a3 and p5 on growth performance, apparent total tract digestibility of nutrients, fecal and intestinal microflora and intestinal morphology in weanling pigs. Livestock Science. 2014;159:53-60.
[48] Yoon JH, Ingale SL, Jin SK, Kim KH, Lohakare J, Park YK, et al. Effects of dietary supplementation with antimicrobial peptide-p5 on growth performance, apparent total tract digestibility, faecal and intestinal microflora and intestinal morphology of weanling pigs. Journal of the Science of Food & Agriculture. 2012;159:53-60.
[49] Liu T, She R, Wang K, Bao H, Zhang Y, Luo D, et al. Effects of rabbit sacculus rotundus antimicrobial peptides on the intestinal mucosal immunity in chickens. Poultry Science. 2008;87:250-4.
[50] Jayaraman S. Bacillus subtilis pb6 improves intestinal health of broiler chickens challenged with clostridium perfringens-induced necrotic enteritis. Poult Sci. 2013;92:370-4.
[51] Xiao H, Tan BE, Wu MM, Yin YL, Li TJ, Yuan DX, et al. Effects of composite antimicrobial peptides in weanling piglets challenged with deoxynivalenol: Ii. Intestinal morphology and function. Journal of Animal Science. 2013;91:4750.
[52] Hu F, Gao X, She R, Chen J, Mao J, Xiao P, et al. Effects of antimicrobial peptides on growth performance and small intestinal function in broilers under chronic heat stress. Poultry Science. 2016:pew379.
[53] Mansour SC, Pena OM, Hancock RE. Host defense peptides: Front-line immunomodulators: Trends in immunology. Trends in Immunology. 2014;35:443.
[54] Hancock REW, Sahl HG. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nature Biotechnology. 2006;24:1551-7.
[55] Lai Y, Gallo RL. Amped up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends in Immunology. 2009;30:131.
[56] Wang D, Ma W, She R, Sun Q, Liu Y, Hu Y, et al. Effects of swine gut antimicrobial peptides on the intestinal mucosal immunity in specific-pathogen-free chickens. Poultry Science. 2009;88:967-74.
[57] Scott MG. The role of antimicrobial peptides in animal defenses. Proceedings of the National Academy of Sciences of the United States of America. 2000;97:8856.
[58] Nijnik A, Lma M. Synthetic cationic peptide idr-1002 provides protection against bacterial infections through chemokine induction and enhanced leukocyte recruitment. Journal of Immunology. 2010;184:2539.
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
27
_____________________________________________________________________________

[59] Hancock REW, Nijnik A, Philpott DJ. Modulating immunity as a therapy for bacterial infections. Nature Reviews Microbiology. 2012;10:243.
[60] Ren ZH, Yuan W, Deng HD, Deng JL, Dan QX, Jin HT, et al. Effects of antibacterial peptide on cellular immunity in weaned piglets. Journal of Animal Science. 2015;93:127-34.
[61] Dijk AV, Eldik MV, Veldhuizen EJA, Bokhoven HLMT, Zoete MRD, Bikker FJ, et al. Immunomodulatory and anti-inflammatory activities of chicken cathelicidin-2 derived peptides. Plos One. 2016;11:e0147919.
[62] Kim WH, Lillehoj HS, Min W. Evaluation of the immunomodulatory activity of the chicken nk-lysin-derived peptide cnk-2. Scientific Reports. 2017;7:45099.
[63] Jin Z, Yang YX, Choi JY, Shinde PL, Yoon SY, Hahn TW, et al. Potato (solanum tuberosum l. Cv. Gogu valley) protein as a novel antimicrobial agent in weanling pigs. Journal of Animal Science. 2008;86:1562-72.
[64] Gläser R, Becker K, Eiff CV, Meyer-Hoffert U, Harder J. Decreased susceptibility of staphylococcus aureus small-colony variants toward human antimicrobial peptides. Journal of Investigative Dermatology. 2014;134:2347-50.
Antimicrobial research: Novel bioknowledge and educational programs (A. Méndez-Vilas, Ed.)
28
_____________________________________________________________________________
Related Documents











